С того момента, как Земля стала полноценной планетой, на ней постепенно начала зарождаться жизнь. Прежде всего, появились клетки - строительный материал для всего живого. Наблюдающееся сейчас разнообразие жизненных форм не только радует глаз, но и заставляет задуматься над сложностью эволюционных процессов. Несмотря на непохожесть организмов, клетки, из которых они состоят, походят друг на друга. Но ученые все же нашли различия и поделили их на две обширные группы: прокариоты и эукариоты. При этом вирусы классифицируются отдельно, а наука пока не может определиться, живые ли это организмы.
Микромир среди нас
Сейчас люди живут в большом мире, зная, что рядом находится маленькая вселенная, кишащая различными микроорганизмами: некоторые из них лояльны к человеку, другие - представляют опасность.
Речь идет о многообразии бактериальной жизни. К прокариотам относятся именно бактерии. Тогда как более сложные эукариоты схематически подразделяются на несколько царств: растения, животные, грибы и слизевики.
Бактерии (прокариоты) отличаются следующими признаками:
- единичная, безъядерная клетка (образование нуклеоида);
- бинарное деление;
- различие форм, величин;
- наличие или отсутствие органелл.
Бактерии-прокариоты подразделяются на извитые (вибрионы), палочковидные (бациллы), похожие на штопор (спириллы) и шарообразные (кокки). Передвигаются клетки либо с помощью слизи, либо используя жгутики.
Флагман эволюции
Доказано, что прокариоты появились первыми. Если перевести слово с греческого языка, то "прокариоты" - это клетки, в которых отсутствует ядро.

Но в столь категоричное определение необходимо внести некоторые поправки. Прокариоты содержат структуры, подобные ядру, не имеющие мембран, которые находятся в цитоплазматической жидкости. Такие неоформленные ядра получили название нуклеоиды. Именно в них находится скромное количество генетической информации клетки.
Возникли простейшие клетки около 35 миллионов веков тому назад. Следуя этой фантастической цифре, можно утверждать, что прокариоты - это первая живая материя на тогда еще юной Земле.
Больше. Объемнее. Сложнее
Обратившись к знаниям древних греков, можно выяснить, что "эукариот" означает «обладающий ядром», и это утверждение полностью соответствует действительности.

Такие клетки, выражаясь современным языком, более продвинуты. Наличие полноценного ядра играет в этом не последнюю роль. Полностью оформленное ядро двумя слоями мембраны надежно защищает генетический код, содержащийся в нем. Частично молекулы ДНК находятся за пределами ядра, в хлоропластах и митохондриях. Кроме прочего, эукариоты значительно больше безъядерных по размеру и объему.
Появились эукариоты примерно спустя два миллиарда лет, после возникновения элементарных прокариот. Можно сказать, что к прокариотам относятся, словно к прародителям истинно ядерных.
Вооружившись микроскопом
Клетки прокариот не только гораздо меньше, нежели эукариоты, они значительно разнятся своими структурными особенностями.

Прокариоты и эукариоты отличаются неодинаковой сложностью строения, где прокариотическая клетка гораздо проще.
- Слизистая капсула. Функции капсулы сводятся к защите клетки: она предохраняет от высыхания и действия антител. Состоит в основном из полисахаридов, с полипептидными включениями.
- Стенка клетки. Благодаря муреину, из которого состоит данная оболочка, этот структурный компонент отличается жесткостью. Выполняет тройную функцию: защитную, транспортную, надежно фиксирует форму и целостность клетки.
- Клеточный протопласт. Из-за разности в росте протопласта и наружной клеточной мембраны случаются инвагинации - внутренние прогибы. Такие процессы позволяют образовывать органоиды, называемые мезосомами. Они выполняют необходимые клеточные функции. В то время как на рибосомы возложена миссия синтезирования белка.
- Нуклеоид. Вытянутая, эллипсоидная структура, выполняющая роль ядра. Она никак не отделена от цитоплазмы, поэтому ученые не классифицируют ее как полноценное ядро. Дополнительную ДНК-информацию могут нести очень мелкие частицы - плазмиды.
- Необязательные органеллы. Представлены различными вкраплениями: серы, полифосфатов, масла, гликогена, полисахаридных зерен. Также к органоидам относят жгутики, которые помогают клетке двигаться, и пили - белковые включения, выполняющие функцию прикрепления.
Если можно говорить о простоте строения применимо к такой сложной живой конструкции, как клетка, то, безусловно, прокариоты - это простые структуры.
Строение истинно ядерных

Эукариоты, напротив, имеют сложное строение. Возникшие, по предположениям ученого мира, на основе прокариот, они содержат в своем составе более совершенные структуры.
- Мембрана. Выполняет избирательно-защитную функцию.
- Клеточная стенка, или гликоликс (у животных). У грибов и растений она играет стабилизирующую и сохраняющую форму роль. Состоит из хитина и целлюлозных волокон, соответственно. Животные обходятся без клеточной стенки по причине действия иных механизмов.
- Цитоплазма. Состоит из раствора неорганических и органических веществ под общим названием гиалоплазма. Главный компонент, способный осуществлять химические реакции, - это белок.
- Ядро. Самая важная составляющая клетки. Оно хранит генетическую информацию в хромосомах. Защищено ядро двумя мембранами.
- Органоиды. Находятся на территории гиалоплазмы и представлены множеством наименований. Наиважнейшие составляющие: митохондрии, комплекс Гольджи, хлоропласты, лизосомы, рибосомы.
Ядро эукариот может делиться двумя способами: митозом и мейозом. Это чрезвычайно важно для развития и работы клетки.
Краткий анализ
Отличия двух групп можно систематизировать и описать на языке, понятном для всех. Тем более что клетки прокариот и эукариот очевидно разнятся.
| Признаки | Прокариотические клетки | Эукариотические клетки |
| Хронологическая ниша | 3,5 миллиарда | полтора миллиарда лет |
| Величина | 0,01 мм | 0,1 мм (больше в 1000 раз) |
| Геном | находится в нуклеоиде, оформленном в виде эллипса | расположен в ядре, содержащем хромосомы; частично ДНК расположена в митохондриях и хлоропластах |
| Органоиды | иногда встречаются, но в малом количестве и безмембранные | во множестве; пластиды и митохондрии имеют свой геном и способность к размножению; каждая органелла выполняет четко определенную функцию |
| Способ деления | бинарный (пополам) | митоз: деление с сохранностью численности хромосом; мейоз: двоекратное сокращение числа хромосом |
| Движение | иногда двигаться помогает слизь, чаще - белковые нити | с помощью жгутиков (сложных, многослойных отростков) и ресничек |
| Явление фагоцитоза | отсутствует | предположительно появилось для обеспечения жизнедеятельности крупной клетки; благодаря этому явлению появились хищники |
Особенности прокариотов
После того как появился электронный микроскоп, стало возможным дифференцировать эукариоты и прокариоты, а также изучить их подробнее.

Главное отличие доядерных клеток в том, что прокариоты - это бактерии, которые присутствуют повсеместно: в организме человека, воде, воздухе, растениях. Очень много бактерий находится в почве. Они живут даже в атомных реакторах!
Несмотря на мелкие размеры, прокариоты являются хранителями закодированной генетической информации, влияющей на жизнедеятельность организмов. Например, в плазмидах, которые находятся в свободном плавании среди вод цитоплазматического коктейля, может содержаться ген, влияющий на устойчивость бактерии к медикаментам.
Кроме того, простота организации не мешает бактериям процветать, активно множась, наряду с более сложными эукариотами.
Дилемма: польза или вред
Значение эукариотов не подвергается сомнениям: благодаря им появились разнообразные формы жизни. А вот с бактериями не все так однозначно. С одной стороны, благодаря микроорганизмам, находящимся в кишечнике, люди могут успешно переваривать пищу, синтезировать и усваивать витамины. Также прокариоты, покрывающие кожу или слизистые человека, выполняют защитную функцию.
Некоторые бактерии необходимы для производства сыров, сметаны, кислой капусты. Интересно, что выделили бактерии, которые обладают действием, сходным с антибиотиками.
С другой стороны, определенные виды безъядерных могут вызывать болезни или портить продукты. Они способны не только выделять токсины, но и разрушать клетки оккупированного организма.
Поэтому наверняка говорить о пользе или вреде прокариотов нельзя: все зависит от их типов. Но в любом случае этим мельчайшим клеткам человечество обязано своим существованием.
Все живые организмы на Земле делятся на две группы: прокариот и эукариот.
- Эукариоты – это растения, животные и грибы.
- Прокариоты – это бактерии (в том числе цианобактерии, они же "сине-зеленые водоросли").
Главное отличие
У прокариот нет ядра , кольцевая ДНК (кольцевая хромосома) расположена прямо в цитоплазме (этот участок цитоплазмы называется нуклеоид).
У эукариот есть оформленное ядро
(наследственная информация [ДНК] отделена от цитоплазмы ядерной оболочкой).
Дополнительные отличия
1) Раз у прокариот нет ядра, то нет и митоза/мейоза. Бактерии размножаются делением надвое ("прямым" делением, в отличие от "непрямого" – митоза).
2) У прокариот рибосомы мелкие (70S), а у эукариот – крупные (80S).
3) У эукариот имеется множество органоидов: митохондрии, эндоплазматическая сеть, клеточный центр, и т.д. Вместо мембранных органоидов у прокариот есть мезосомы – выросты плазматической мембраны, похожие на кристы митохондрий.
4) Клетка прокариот гораздо меньше клетки эукариот: по диаметру в 10 раз, по объему - в 1000 раз.
Сходство
Клетки всех живых организмов (всех царств живой природы) содержат плазматическую мембрану, цитоплазму и рибосомы.
Выберите один, наиболее правильный вариант. ВЫБЕРИТЕ НЕВЕРНОЕ УТВЕРЖДЕНИЕ. У бактерий отсутствуют
1) половые клетки
2) мейоз и оплодотворение
3) митохондрии и клеточный центр
4) цитоплазма и ядерное вещество
Ответ
Выберите один, наиболее правильный вариант. В прокариотических клетках реакции окисления происходят на
1) рибосомах в цитоплазме
2) впячиваниях плазматической мембраны
3) оболочках клеток
4) кольцевой молекуле ДНК
Ответ
ПРОКАРИОТЫ
1. Выберите три варианта. Бактериальную клетку относят к группе прокариотических, так как она
1) не имеет ядра, покрытого оболочкой
2) имеет цитоплазму
3) имеет одну молекулу ДНК, погруженную в цитоплазму
4) имеет наружную плазматическую мембрану
5) не имеет митохондрий
6) имеет рибосомы, где происходит биосинтез белка
Ответ
2. Выберите три варианта. Почему бактерии относят к прокариотам?
1) содержат в клетке ядро, обособленное от цитоплазмы
2) состоят из множества дифференцированных клеток
3) имеют одну кольцевую хромосому
4) не имеют клеточного центра, комплекса Гольджи и митохондрий
5) не имеют обособленного от цитоплазмы ядра
6) имеют цитоплазму и плазматическую мембрану
Ответ
3. Выберите три варианта. Для прокариотной клетки характерно наличие
1) рибосом
2) митохондрий
3) оформленного ядра
4) плазматической мембраны
5) эндоплазматической сети
6) одной кольцевой ДНК
Ответ
4. Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. У прокариотических клеток есть
1) нуклеоид с ДНК
2) настоящее ядро
3) аппарат Гольджи
4) гомологичные хромосомы
5) рибосомы
6) клеточная мембрана
Ответ
СОБИРАЕМ 5:
А) отсутствие мембранных органоидов
Б) отсутствие рибосом в цитоплазме
В) образование двух и более хромосом линейной структуры
ПРОКАРИОТЫ КРОМЕ
1. Все перечисленные признаки, кроме двух, используются для описания прокариотической клетки. Определите два признака, «выпадающие» из общего списка, и запишите цифры, под которыми они указаны.
1) Отсутствие в ней оформленного ядра
2) Наличие цитоплазмы
3) Наличие клеточной мембраны
4) Наличие митохондрий
5) Наличие эндоплазматической сети
Ответ
2. Все перечисленные ниже признаки, кроме двух, характеризуют строение бактериальной клетки. Определите два признака, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) отсутствие оформленного ядра
2) наличие лизосом
3) наличие плотной оболочки
4) отсутствие митохондрий
5) отсутствие рибосом
Ответ
3. Перечисленные ниже понятия, кроме двух, используются для характеристики прокариот. Определите два понятия, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) митоз
2) спора
3) гамета
4) нуклеоид
5) мезосома
Ответ
4. Все приведенные ниже термины, кроме двух, используются для описания строения бактериальной клетки. Определите два термина, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) неподвижная цитоплазма
2) кольцевая молекула ДНК
3) мелкие (70S) рибосомы
4) способность к фагоцитозу
5) наличие ЭПС
Ответ
5. Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Прокариотическая клетка не имеет в своем составе
1) ядра
2) цитоплазмы
3) эндоплазматической сети
4) плазматической мембраны
5) рибосом
6) пластид
Ответ
ПРОКАРИОТЫ - ЭУКАРИОТЫ
1. Установите соответствие между характеристикой клетки и типом организации этой клетки: 1) прокариотический, 2) эукариотический
А) клеточный центр участвует в образовании веретена деления
Б) в цитоплазме находятся лизосомы
В) хромосома образована кольцевой ДНК
Г) отсутствуют мембранные органоиды
Д) клетка делится митозом
Е) мембрана образует мезосомы
Ответ
2. Установите соответствие между характеристикой клетки и её типом: 1) прокариотическая, 2) эукариотическая
А) мембранные органоиды отсутствуют
Б) имеется клеточная стенка из муреина
В) наследственный материал представлен нуклеоидом
Г) содержит только мелкие рибосомы
Д) наследственный материал представлен линейными ДНК
Е) клеточное дыхание происходит в митохондриях
Ответ
3. Установите соответствие между признаком и группой организмов: 1) Прокариоты, 2) Эукариоты. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) отсутствие ядра
Б) наличие митохондрий
В) отсутствие ЭПС
Г) наличие аппарата Гольджи
Д) наличие лизосом
Е) линейные хромосомы, состоящие из ДНК и белка
Ответ
4. Установите соответствие между органоидами и клетками, которые их имеют: 1) прокариотическая, 2) эукариотическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) аппарат Гольджи
Б) лизосомы
В) мезосомы
Г) митохондрии
Д) нуклеоид
Е) ЭПС
Ответ
5. Установите соответствие между клетками и их особенностями: 1) прокариотическая, 2) эукариотическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) молекула ДНК кольцевая
Б) поглощение веществ путем фаго- и пиноцитоза
В) образуют гаметы
Г) рибосомы мелкие
Д) есть мембранные органоиды
Е) характерно прямое деление
Ответ
6. Установите соответствие между клетками и их особенностями: 1) прокариотическая, 2) эукариотическая. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
1) наличие обособленного ядра
2) образование спор для перенесения неблагоприятных условий среды
3) расположение наследственного материала только в замкнутой ДНК
4) деление путем мейоза
5) способность к фагоцитозу
6) отсутствуют митохондрии
Ответ
Установите соответствие между характеристиками и типами клеток, изображенных на рисунке. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) имеют мезосомы
Б) осмотрофный способ питания
В) делятся митозом
Г) имеют развитую ЭПС
Д) образуют споры при неблагоприятных условиях
Е) имеют оболочку из муреина
Ответ
ПРОКАРИОТЫ - ЭУКАРИОТЫ ОТЛИЧИЯ
1. Выберите три варианта. Клетки прокариот отличаются от клеток эукариот
1) наличием нуклеоида в цитоплазме
2) наличием рибосом в цитоплазме
3) синтезом АТФ в митохондриях
4) присутствием эндоплазматической сети
5) отсутствием морфологически обособленного ядра
6) наличием впячиваний плазматической мембраны, выполняющих функцию мембранных органоидов
Ответ
2. Выберите три варианта. Прокариотные клетки отличаются от эукариотных
1) наличием рибосом
2) отсутствием митохондрий
3) отсутствием оформленного ядра
4) наличием плазматической мембраны
5) отсутствием органоидов движения
6) наличием одной кольцевой хромосомы
Ответ
БАКТЕРИИ - РАСТЕНИЯ ПРИЗНАКИ
1. Установите соответствие между признаком и царством: 1) бактерии, 2) растения. Запишите цифры 1 и 2 в правильном порядке.
A) все представители прокариоты
Б) все представители эукариоты
B) могут делиться пополам
Г) есть ткани и органы
Д) есть фото и хемосинтетики
Е) хемосинтетики не встречаются
Ответ
2. Установите соответствие между признаками организмов и их царством: 1) бактерии, 2) растения. Запишите цифры 1 и 2 в правильном порядке.
А) различные представители способны к фотосинтезу и хемосинтезу
Б) в наземных экосистемах превосходят все другие группы по биомассе
В) клетки делятся путем митоза и мейоза
Г) имеют пластиды
Д) клеточные стенки обычно не содержат целлюлозы
Е) лишены митохондрий
Ответ
БАКТЕРИИ - РАСТЕНИЯ СХОДСТВО
1. Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Сходство бактерий и растений состоит в том, что они
1) прокариотические организмы
2) образуют споры при неблагоприятных условиях
3) имеют клеточное тело
4) среди них имеются автотрофы
5) обладают раздражимостью
6) способны к вегетативному размножению
Ответ
2. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны. Сходство клеток бактерий и растений состоит в том, что они имеют
1) рибосомы
2) плазматическую мембрану
3) оформленное ядро
4) клеточную стенку
5) вакуоли с клеточным соком
6) митохондрии
Ответ
БАКТЕРИИ - РАСТЕНИЯ ОТЛИЧИЯ
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Бактерии, в отличие от низших растений,
1) по типу питания бывают хемотрофами
2) при размножении образуют зооспоры
3) не имеют мембранных органоидов
4) имеют слоевище (таллом)
5) при неблагоприятных условиях образуют споры
6) синтезируют полипептиды на рибосомах
Ответ
БАКТЕРИИ - ЖИВОТНЫЕ ПРИЗНАКИ
1. Установите соответствие между признаком и организмом, для которого он характерен: 1) инфузория туфелька, 2) бацилла сибирской язвы. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) клетка не имеет ядерной мембраны
Б) образует споры вне организма хозяина
В) клетка содержит оформленное ядро
Г) не имеет аппарата Гольджи
Д) передвигается с помощью ресничек
Е) имеется пищеварительная вакуоль
Ответ
2. Установите соответствие между органоидами и их наличием у клеток: 1) бактерий, 2) животных. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) лизосомы
Б) клеточная стенка
В) ядрышко
Г) аппарат Гольджи
Д) кольцевая ДНК
Е) мезосомы
Ответ
БАКТЕРИИ - ЖИВОТНЫЕ СХОДСТВО
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Сходство клеток животных и бактерий состоит в том, что они имеют
1) рибосомы
2) цитоплазму
3) гликокаликс
4) митохондрии
5) оформленное ядро
6) цитоплазматическую мембрану
Ответ
БАКТЕРИИ - ГРИБЫ ПРИЗНАКИ
1. Установите соответствие между признаком организма и царством, для которого он характерен: 1) грибы, 2) бактерии
А) ДНК замкнута в виде кольца
Б) по способу питания - автотрофы или гетеротрофы
В) клетки имеют оформленное ядро
Г) ДНК имеет линейное строение
Д) в клеточной стенке имеется хитин
Е) ядерное вещество расположено в цитоплазме
Ответ
2. Установите соответствие между признаками организмов и царствами, для которых они характерны: 1) Грибы, 2) Бактерии. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) образование микоризы с корнями высших растений
Б) образование клеточной стенки из хитина
В) тело в виде мицелия
Г) размножение спорами
Д) способность к хемосинтезу
Е) расположение кольцевой ДНК в нуклеоиде
Ответ
3. Установите соответствие между характеристиками и организмами: 1) дрожжи, 2) кишечная палочка. Запишите цифры 1 и 2 в порядке, соответствующем буквам.
А) геном представлен одной кольцевой молекулой ДНК
Б) клетка покрыта оболочкой из муреина
В) делится митозом
Г) в анаэробных условиях вырабатывает этанол
Д) имеет жгутики
Е) не имеет мембранных органоидов
Ответ
БАКТЕРИИ - ГРИБЫ СХОДСТВА
Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. Бактерии, как и грибы,
1) составляют особое царство
2) являются только одноклеточными организмами
3) размножаются с помощью спор
4) являются редуцентами в экосистеме
5) могут вступать в симбиоз
6) поглощают вещества из почвы с помощью гифов
Ответ
БАКТЕРИИ - ГРИБЫ ОТЛИЧИЯ
Выберите три варианта. Бактерии, в отличие от шляпочных грибов,
1) одноклеточные организмы
2) многоклеточные организмы
3) имеют в клетках рибосомы
4) не имеют митохондрий
5) доядерные организмы
6) не имеют цитоплазмы
Ответ
БАКТЕРИИ - ГРИБЫ - РАСТЕНИЯ
Установите соответствие между характеристикой клетки и ее типом: 1) бактериальная, 2) грибная, 3) растительная. Напишите цифры 1, 2 и 3 в правильной последовательности.
А) отсутствие мембранных органоидов
Б) запасающее вещество – крахмал
В) способность к хемосинтезу
Г) наличие нуклеоида
Д) наличие хитина в клеточной стенке
Ответ
БАКТЕРИИ - ЖИВОТНЫЕ - РАСТЕНИЯ
Установите соответствие между признаками клеток и царством: 1) Растения, 2) Животные, 3) Бактерии. Запишите цифры 1-3 в порядке, соответствующем буквам.
А) включает только одноклеточные организмы
Б) клетки безъядерные
В) фотосинтез проходит в хлоропластах
Г) клетки не имеют клеточных стенок
Д) запасным веществом клеток является гликоген
Е) группы клеток образуют механические и проводящие ткани
Ответ
ГРИБЫ - БАКТЕРИИ ОТЛИЧИЯ
Выберите три варианта. Чем отличаются грибы от бактерий?
1) составляют группу ядерных организмов (эукариот)
2) относятся к гетеротрофным организмам
3) размножаются спорами
4) одноклеточные и многоклеточные организмы
5) при дыхании используют кислород воздуха
6) участвуют в круговороте веществ в экосистеме
Ответ
Ответ
ЭУКАРИОТЫ - ПРОКАРИОТЫ ОТЛИЧИЯ
Выберите три варианта. Клетки эукариотных организмов, в отличие от прокариотных, имеют
1) цитоплазму
2) ядро, покрытое оболочкой
3) молекулы ДНК
4) митохондрии
5) плотную оболочку
6) эндоплазматическую сеть
Ответ

Проанализируйте таблицу. Заполните пустые ячейки таблицы, используя понятия и термины, приведенные в списке.
1) митоз, мейоз
2) перенесение неблагоприятных условий среды
3) перенос информации о первичной структуре белка
4) двумембранные органоиды
5) шероховатая эндоплазматическая сеть
6) мелкие рибосомы
Ответ

Выберите три верных ответа из шести и запишите цифры, под которыми они указаны. В процессе эволюции сформировались организмы разных царств. Какие признаки характерны для царства, представитель которого изображен на рисунке.
1) клеточная стенка состоит в основном из муреина
2) хроматин содержится в ядрышке
3) хорошо развита эндоплазматическая сеть
4) отсутствуют митохондрии
5) наследственная информация содержится в кольцевой молекуле ДНК
6) пищеварение происходит в лизосомах
Ответ

1. Все перечисленные ниже термины, кроме двух, используют для описания клетки, изображённой на рисунке. Определите два термина, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) замкнутая молекула ДНК
2) мезосома
3) мембранные органоиды
4) клеточный центр
5) нуклеоид
Ответ
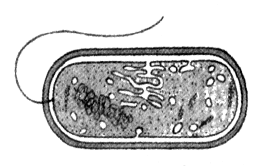
2. Все перечисленные ниже признаки, кроме двух, используются для описания изображенной на рисунке клетки. Определите два термина, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) деление путем митоза
2) наличие клеточной стенки из муреина
3) наличие нуклеоида
4) отсутствие мембранных органоидов
5) поглощение веществ путем фаго- и пиноцитоза
Ответ

3. Все перечисленные ниже термины, кроме двух, используют для описания клетки, изображённой на рисунке. Определите два термина, «выпадающих» из общего списка, и запишите цифры, под которыми они указаны.
1) замкнутая ДНК
2) митоз
3) гаметы
4) рибосомы
5) нуклеоид
Ответ
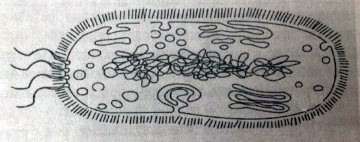
4. Все перечисленные ниже признаки, кроме двух, можно использовать для описания изображённой на рисунке клетки. Определите два признака, «выпадающих» из общего списка и запишите цифры, под которыми они указаны.
1) есть клеточная мембрана
2) есть аппарат Гольджи
3) есть несколько линейных хромосом
4) есть рибосомы
5) есть клеточная стенка
Ответ

5сб. Все перечисленные ниже признаки, кроме двух, можно использовать для описания изображённой на рисунке клетки. Определите два признака, «выпадающих» из общего списка и запишите цифры, под которыми они указаны.
1) имеют линейные хромосомы
2) характерно бинарное деление
3) имеет эндоплазматическую сеть
4) образует спору
5) содержит мелкие рибосомы
Ответ
СОБИРАЕМ 6:
1) плазмида
2) дыхание в митохондриях
3) деление надвое
Все перечисленные ниже признаки, кроме двух, можно использовать для описания прокариотической ДНК. Определите два признака, выпадающих из общего списка, и запишите цифры, под которыми они указаны.
1) содержит аденин, гуанин, урацил и цитозин
2) состоит из двух цепей
3) имеет линейную структуру
4) не связана со структурными белками
5) лежит в цитоплазме
Ответ
© Д.В.Поздняков, 2009-2019
Появление эукариот – важнейшее событие. Изменило структуру биосферы и открыло принципиально новые возможности для прогрессивной эволюции. Эукариотическая клетка является результатом долгой эволюции мира прокариот, мира, в котором разнообразные микробы приспосабливались друг к другу и искали способы эффективной кооперации.
набросок хронологии (повторение)
Фотосинтезирующий прокариотический комплекс Chlorochromatium aggregatum.
Эукариоты возникли в результате симбиоза нескольких видов прокариот. Прокариоты вообще весьма склонны к симбиозу (см. главу 3 в книге «Рождение сложности»). Вот интересная симбиотическая система, известная под названием Chlorochromatium aggregatum. Живет в глубоких озерах, где есть на глубине бескислородные условия. Центральный компонент – подвижная гетеротрофная бета-протеобактерия. Вокруг нее стопками располагаются от 10 до 60 фотосинтезирующих зеленых серных бактерий. Все компоненты соединены выростами наружной мембраны центральной бактерии. Смысл содружества в том, что подвижная бета-протеобактерия перетаскивает всю компанию в места, благоприятные для жизни привередливых серных бактерий, а серные бактерии занимаются фотосинтезом и обеспечивают пищей и себя, и бета-протеобактерию. Может быть, какие-то древние микробные ассоциации примерно такого типа и были предками эукариот.
Теория симбиогенеза. Мережковский, Маргулис. Митохондрии – потомки альфа-протеобактерий, пластиды – потомки цианобактерий. Труднее понять, кто был предком всего остального, то есть цитоплазмы и ядра. Ядро и цитоплазма эукариот сочетает в себе признаки архей и бактерий, а также имеет множество уникальных особенностей.
О митохондриях. Возможно, именно приобретение митохондрий (а не ядра) было ключевым моментом в становлении эукариот. Большинство генов предков митохондрий было перенесено в ядро, где они попали под контроль ядерных регуляторных систем. Эти ядерные гены митохондриального происхождения кодируют не только белки митохондрий, но и многие белки, работающие в цитоплазме. Это говорит о том, что митохондриальный симбионт сыграл более важную роль в формировании эукариотической клетки, чем предполагалось.
Совместное существование в одной клетке двух разных геномов требовало развития эффективной системы их регуляции. А для того, чтобы эффективно управлять работой большого генома, необходимо изолировать геном от цитоплазмы, в которой протекает обмен веществ и идут тысячи химических реакций. Ядерная оболочка как раз и отделяет геном от бурных химических процессов цитоплазмы. Приобретение симбионтов (митохондрий) могло стать важным стимулом в развитии ядра и генно-регуляторных систем.
То же относится и к половому размножению. Без полового размножения можно жить до тех пор, пока геном у вас достаточно маленький. Организмы с большим геномом, но лишенные полового размножения, обречены на быстрое вымирание, за редчайшими исключениями.
Альфапротеобактерии – к этой группе относились предки митохондрий.
Родоспириллум – удивительный микроорганизм, который может жить и за счет фотосинтеза, в том числе и в анаэробных условиях, и как аэробный гетеротроф, и даже как аэробный хемоавтотроф. Он может, например, расти за счет окисления угарного газа СО, не используя никаких других источников энергии. Вдобавок ко всему этому, он умеет еще и фиксировать атмосферный азот. То есть это в высшей степени универсальный организм.
Иммунная система принимает митохондрии за бактерии. Когда при травме в кровь попадают разрушенные митохондрии, из них высвобождаются характерные молекулы, которые встречаются только у бактерий и у митохондрий (кольцевая ДНК бактериального типа и белки, несущие на одном из своих концов особую модифицированную аминокислоту формилметионин). Это связано с тем, что аппарат синтеза белка в митохондриях остался таким же, как у бактерий. Клетки иммунной системы – нейтрофилы – реагируют на эти митохондриальные вещества точно так же, как на бактериальные, и при помощи тех же рецепторов. Это ярчайшее подтверждение бактериальной природы митохондрий.
Главная функция митохондрий – кислородное дыхание. Скорее всего, стимулом для объединения анаэробного предка ядра и цитоплазмы с «протомитохондрией» была необходимость защититься от токсического действия кислорода.
Откуда взялись у бактерий, и в том числе у альфапротеобактерий, молекулярные системы, необходимые для кислородного дыхания? Похоже, в их основу были положены молекулярные системы фотосинтеза. Электронно-транспортная цепь, сформировавшаяся у бактерий как часть фотосинтетического аппарата, была адаптирована для кислородного дыхания. У некоторых бактерий до сих пор участки электронно-транспортных цепей используются одновременно и в фотосинтезе, и в дыхании. Скорее всего предками митохондрий были аэробные гетеротрофные альфа-протеобактерии, которые, в свою очередь, произошли от фотосинтезирующих альфа-протеобактерий, таких как родоспириллум.
Число общих и уникальных белковых доменов у архей, бактерий и эукариот. Белковый домен – это часть белковой молекулы, имеющая определенную функцию и характерную структуру, то есть последовательность аминокислот. Каждый белок, как правило, содержит один или несколько таких структурно-функциональных блоков, или доменов.
4,5 тысячи белковых доменов, которые есть у эукариот, можно разделить на 4 группы: 1) имеющиеся только у эукариот, 2) общие для всех трех надцарств, 3) общие для эукариот и бактерий, но отсутствующие у архей; 4) общие для эукариот и архей, но отсутствующие у бактерий. Мы рассмотрим две последние группы (они на рисунке выделены цветом), поскольку для этих белков можно с определенной уверенностью говорить об их происхождении: соответственно бактериальном или архейном.
Ключевой момент в том, что эукариотические домены, предположительно унаследованные от бактерий и от архей, имеют существенно разные функции. Домены, унаследованные от архей (их функциональный спектр показан на левом графике), играют ключевую роль в жизни эукариотической клетки. Среди них преобладают домены, связанные с хранением, воспроизведением, организацией и считыванием генетической информации. Большинство "архейных" доменов относится к тем функциональным группам, в пределах которых горизонтальный обмен генами у прокариот происходит реже всего. Видимо, эукариоты получили этот комплекс путем прямого (вертикального) наследования от архей.
Среди доменов бактериального происхождения тоже есть белки, связанные с информационными процессами, но их мало. Большинство из них работает только в митохондриях или пластидах. Эукариотические рибосомы цитоплазмы имеют архейное происхождение, рибосомы митохондрий и пластид имеют бактериальное происхождение.
Среди бактериальных доменов эукариот значительно выше доля сигнально-регуляторных белков. От бактерий эукариоты унаследовали многие белки, ответственные за механизмы реагирования клетки на факторы внешней среды. А также – многие белки, связанные с обменом веществ (подробнее см. в главе 3 «Рождения сложности»).
Эукариоты имеют:
· Архейную «сердцевину» (механизмы работы с генетической информацией и синтеза белка)
· Бактериальную «периферию» (обмен веществ и сигнально-регуляторные системы)
· Простейший сценарий: АРХЕЯ проглотила БАКТЕРИЙ (предков митохондрий и пластид) и все свои бактериальные признаки приобрела от них.
· Этот сценарий слишком прост, потому что у эукариот много бактериальных белков, которые не могли быть заимствованы у предков митохондрий или пластид.
У эукариот много «бактериальных» доменов, не характерных ни для цианобактерий (предков пластид), ни для альфапротеобактерий (предков митохондрий). Они были получены от каких-то других бактерий.
Птицы и динозавры. Реконструировать прото-эукариот трудно. Ясно, что та группа древних прокариот, которая дала начало ядру и цитоплазме, обладала рядом уникальных особенностей, которых нет у прокариот, доживших до наших дней. И когда мы пытаемся реконструировать облик этого предка, мы сталкиваемся с тем, что простор для гипотез оказывается слишком большим.
Аналогия. Известно, что птицы произошли от динозавров, причем не от каких-то неизвестных динозавров, а от вполне определенной группы – манирапторых динозавров, которые относятся к тероподам, а тероподы в свою очередь – это одна из групп ящеротазовых динозавров. Найдено много переходных форм между нелетающими динозаврами и птицами.
Но что бы мы могли сказать о предках птиц, если бы ископаемой летописи не было? В лучшем случае мы бы выяснили, что ближайшей родней птиц являются крокодилы. Но смогли бы мы воссоздать облик прямых предков птиц, то есть динозавров? Вряд ли. Но именно в таком положении мы и находимся, когда пытаемся восстановить облик предка ядра и цитоплазмы. Ясно, что это была группа неких прокариотических динозавров, группа вымершая и не оставившая, в отличие от настоящих динозавров, внятных следов в геологической летописи. Современные археи по отношению к эукариотам – это как современные крокодилы по отношению к птицам. Попробуйте восстановить строение динозавров, зная только птиц и крокодилов.
Аргумент в пользу того, что в докембрии жило много всяких микробов, не похожих на нынешние. Протерозойские строматолиты были намного сложнее и разнообразнее современных. Строматолиты – продукт жизнедеятельности микробных сообществ. Не значит ли это, что и протерозойские микробы были разнообразнее современных, и что многие группы протерозойских микробов просто не дожили до наших дней?
Предковое сообщество эукариот и происхождение эукариотической клетки (возможный сценарий)
Гипотетическое «предковое сообщество» - типичный бактериальный мат, только в его верхнем жили предки цианобактерий, еще не перешедшие к оксигенному фотосинтезу. Они занимались аноксигенным фотосинтезом.Донором электронов служила не вода, а сероводород. В качестве побочного продукта выделялись сера и сульфаты.
Во втором слое обитали пурпурные фотосинтезирующие бактерии, в том числе – альфапротеобактерии, предки митохондрий. Пурпурные бактерии используют длинноволновой свет (красный и инфракрасный). Эти волны обладают лучшей проникающей способностью. Пурпурные бактерии и сейчас часто живут под слоем цианобактерий. Пурпурные альфапротеобактерии тоже используют в качестве донора электрона сероводород.
В третьем слое были бактерии-бродильщики, перерабатывающие органику; некоторые из них в качестве отходов выделяли водород. Это создавало базу для сульфатредуцирующих бактерий. Там могли быть и метаногенные археи. Среди обитавших здесь архей были и предки ядра и цитоплазмы.
Начало кризисным событиям положил переход цианобактерий к кислородному фотосинтезу. В качестве донора электрона цианобактерии начали использовать вместо сероводорода обычную воду. Это открывало большие возможности, но имело и негативные последствия. Вместо серы и сульфатов при фотосинтезе стал выделяться кислород – вещество крайне токсичное для всех древних обитателей земли.
Первыми с этим ядом столкнулись его производители – цианобактерии. Они же, вероятно, первыми стали вырабатывать средства защиты от него. Электронно-транспортные цепи, служившие для фотосинтеза, были модифицированы и начали служить для аэробного дыхания. Изначальная цель, видимо, заключалась не в получении энергии, а только в нейтрализации кислорода.
Вскоре и обитателям второго слоя сообщества – пурпурным бактериям – пришлось вырабатывать аналогичные системы защиты. Так же, как и цианобактерии, они сформировали системы аэробного дыхания на основе фотосинтетических систем. Именно у пурпурных альфапротеобактерий развилась наиболее совершенная дыхательная цепь, которая ныне функционирует в митохондриях эукариот.
В третьем слое сообщества появление свободного кислорода должно было вызвать кризис. Метаногены и многие сульфатредукторы утилизируют молекулярный водород при помощи ферментов-гидрогеназ. Такие микробы не могут жить в аэробных условиях, потому что кислород ингибирует гидрогеназы. Многие бактерии, выделяющие водород, в свою очередь, не растут в среде, где нет микроорганизмов, его утилизирующих. Из бродильщиков в составе сообщества, по-видимому, остались формы, выделяющие в качестве конечных продуктов низкоорганические соединения (пируват, лактат, ацетат и т.п.). Эти бродильщики выработали свои средства защиты от кислорода, менее эвффективные. К числу выживших относились и археи – предки ядра и цитоплазмы.
Может быть, в этот кризисный момент и произошло ключевое событие – ослабление генетической изоляции у предков эукариот и начало активного заимствования чужих генов. Прото-эукариоты инкорпорировали гены разных бродильщиков до тех пор, пока не стали сами микроаэрофильными бродильщиками, сбраживающими углеводы до пирувата и молочной кислоты.
Обитатели третьего слоя – предки эукариот – теперь непосредственно контактировали с новыми обитателями второго слоя – аэробными альфапротеобактериями, которые научились использовать кислород для получения энергии. Метаболизм прото-эукариот и альфапротеобактерий стал взаимодополнительным, что создавало предпосылки для симбиоза. Да и само расположение альфапротеобактерий в сообществе (между верхним, выделяющим кислород, и нижним слоем) предопределяло их роль как «защитников» предков эукариот от избытков кислорода.
Вероятно, прото-эукариоты заглатывали и приобретали в качестве эндосимбионтов многих разных бактерий. Экспериментирование такого рода и сейчас продолжается у одноклеточных эукариот, обладающих огромным разнообразием внутриклеточных симбионтов. Из этих экспериментов союз с аэробными альфапротеобактериями оказался наиболее удачным.
Все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными , но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез - симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу , и поглощенными этой клеткой бактериями - предшественниками митохондрий и пластидов .
Строение эукариотической клетки
См. также категорию Структуры эукариотической клеткиЭукариотические клетки в среднем намного крупнее прокариотических , разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы , что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро - это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК , «упакованные» в хромосомы . Ядро обычно одно, но бывают и многоядерные клетки.
Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных . Затем было выделено царство грибов , которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших , миксомицетов , хромистов . Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона - Unikonta и Bikonta . Положение таких эукариот, как коллодиктион (Collodictyon ) и Diphylleia , на данный момент не определено.
Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки - нуклеоиде , который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды . По своему строению и жизнедеятельности они поразительно похожи на бактерий . Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза , в том числе у многих групп - фагоцитоза . Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники .
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты , грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов , диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина . Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие - строение жгутиков. У бактерий они тонкие - всего 15–20 нм в диаметре. Это полые нити из белка флагеллина . Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
См. также
| Эукариоты в Викисловаре | |
| Эукариоты на Викискладе |
Зарубежная литература
- Bisby FA, Roskov YR, Ruggiero MA, Orrell TM, Paglinawan LE, et al. Species 2000 & ITIS catalogue of life: 2007 annual checklist. Species 2000. Retrieved Jan. 2007. 21, 2008
- Patterson DJ. The diversity of eukaryotes. Am Nat. 1999
- Stechmann A, Cavalier-Smith T. Rooting the eukaryote tree by using a derived gene fusion. Science. 2002
- Richards TA, Cavalier-Smith T. Myosin domain evolution and the primary divergence of eukaryotes. Nature. 2005
- Stechmann A, Cavalier-Smith T. Phylogenetic analysis of eukaryotes using heat-shock protein Hsp90. J Mol Evol. 2003
- Makiuchi T, Nara T, Annoura T, Hashimoto T, Aoki T. Occurrence of multiple, independent gene fusion events for the fifth and sixth enzymes of pyrimidine biosynthesis in different eukaryotic groups. Gene. 2007
- Kim E, Simpson AGB, Graham LE. Evolutionary relationships of apusomonads inferred from taxon-rich analyses of 6 nuclear encoded genes. Mol Biol Evol. 2006
- Nozaki H, Matsuzaki M, Misumi O, Kuroiwa H, Higashiyama T, et al. Phylogenetic implications of the CAD complex from the primitive red alga Cyanidioschyzon merolae (Cyanidiales, Rhodophyta). J Phycol. 2005
- Adl SM, Simpson AGB, Farmer MA, Andersen RA, Anderson OR, et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J Eukaryot Microbiol. 2005
- Keeling PJ, Burger G, Durnford DG, Lang BF, Lee RW, et al. The tree of eukaryotes. Trends Ecol Evol. 2005
- Simpson AGB, Roger AJ. The real ‘kingdoms’ of eukaryotes. Curr Biol. 2004
- Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhattacharya D, et al. Evaluating support for the current classification of eukaryotic diversity. PLoS Genet. 2006
- Burki F, Shalchian-Tabrizi K, Minge M, Skjaeveland A, Nikolaev SI, et al. Phylogenomics reshuffles the eukaryotic supergroups. PLoS ONE. 2007
- Bodyl A. Do plastid-related characters support the chromalveolate hypothesis? J Phycol. 2005
- Stiller JW, Riley J, Hall BD. Are red algae plants? A critical evaluation of three key molecular data sets. J Mol Evol. 2001
- Grzebyk D, Katz ME, Knoll AH, Quigg A, Raven JA, et al. Response to comment on “The evolution of modern eukaryotic phytoplankton”. Science. 2004
- Yoon HS, Grant J, Tekle YI, Wu M, Chaon BC, et al. Broadly sampled multigene trees of eukaryotes. BMC Evol Biol. 2008
- Jarvis P, Soll M. Toc, Tic, and chloroplast protein import. Biochim Biophys Acta. 2001
- Marin B, Nowack ECM, Melkonian M. A plastid in the making: primary endosymbiosis. Protist. 2005
- Nowack ECM, Melkonian M, Glockner G. Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr Biol. 2008
- Theissen U, Martin W. The difference between organelles and endosymbionts. Curr Biol. 2006
- Bhattacharya D, Archibald JM. The difference between organelles and endosymbionts - response to Theissen and Martin. Curr Biol. 2006
- Okamoto N, Inouye I. The katablepharids are a distant sister group of the Cryptophyta: a proposal for Katablepharidophyta divisio nova/Kathablepharida phylum novum based on SSU rDNA and beta-tubulin phylogeny. Protist. 2005
- Andersen RA. Biology and systematics of heterokont and haptophyte algae. Am J Bot. 2004
- Cavalier-Smith T. Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J Eukaryot Microbiol. 1999
- Graham LE, Wilcox LW. Algae. Upper Saddle River, NJ: Prentice Hall; 2000
- Schnepf E, Elbrachter M. Dinophyte chloroplasts and phylogeny: a review. Grana. 1999
- Kohler S, Delwiche CF, Denny PW, Tilney LG, Webster P, et al. A plastid of probable green algal origin in apicomplexan parasites. Science. 1997
- Kohler S. Multi-membrane-bound structures of Apicomplexa: I. the architecture of the Toxoplasma gondii apicoplast. Parasitol Res. 2005
- Hopkins J, Fowler R, Krishna S, Wilson I, Mitchell G, et al. The plastid in Plasmodium falciparum asexual blood stages: a three-dimensional ultrastructural analysis. Protist. 1999
- Tomova C, Geerts WJC, Muller-Reichert T, Entzeroth R, Humbel BM. New comprehension of the apicoplast of Sarcocystis by transmission electron tomography. Biol Cell. 2006
- Moore RB, Obornik M, Janouskovec J, Chrudimsky T, Vancova M, et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature. 2008
- Stiller JW, Reel DC, Johnson JC. A single origin of plastids revisited: convergent evolution in organellar genome content. J Phycol. 2003
- Larkum AWD, Lockhart PJ, Howe CJ. Shopping for plastids. Trends Plant Sci. 2007
- McFadden GI, van Dooren GG. Evolution: red algal genome affirms a common origin of all plastids. Curr Biol. 2004
- Stiller JW, Hall BD. The origin of red algae: implications for plasmid evolution. Proc Natl Acad Sci U S A. 1997
- Sanchez-Puerta MV, Bachvaroff TR, Delwiche CF. Sorting wheat from chaff in multi-gene analyses of chlorophyll c-containing plastids. Mol Phylogenet Evol. 2007
- Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, et al. The evolution of modern eukaryotic phytoplankton. Science. 2004
- Fast NM, Kissinger JC, Roos DS, Keeling PJ. Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol Biol Evol. 2001
- Bucknam J, Boucher Y, Bapteste E. Refuting phylogenetic relationships. Biol Direct. 2006
- Gupta RS, Golding GB. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. J Mol Evol. 1993
- Gupta RS, Singh B. Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr Biol. 1994
- Gomez-Lorenzo MG, Spahn CMT, Agrawal RK, Grassucci RA, Penczek P, et al. Three-dimensional cryo-electron microscopy localization of EF2 in the Saccharomyces cerevisiae 80S ribosome at 17.5 angstrom resolution. EMBO J. 2000
- Jorgensen R, Merrill AR, Andersen GR. The life and death of translation elongation factor 2. Biochem Soc Trans. 2006
- Moreira D, Le Guyader H, Philippe H. The origin of red algae and the evolution of chloroplasts. Nature. 2000
- Germot a, Philippe H. Critical analysis of eukaryotic phylogeny: a case study based on the HSP70 family. J Eukaryot Microbiol. 1999
- Philippe H, Delsuc F, Brinkmann H, Lartillot N. Phylogenomics. Annu Rev Ecol Evol Syst. 2005
- Wiens JJ. Missing data and the design of phylogenetic analyses. J Biomed Inform. 2006
- Philippe H, Snell EA, Bapteste E, Lopez P, Holland PWH, et al. Phylogenomics of eukaryotes: Impact of missing data on large alignments. Mol Biol Evol. 2004
- Patron NJ, Inagaki Y, Keeling PJ. Multiple gene phylogenies support the monophyly of cryptomonad and haptophyte host lineages. Curr Biol. 2007
- Hackett JD, Yoon HS, Li S, Reyes-Prieto A, Rummele SE, et al. Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of Rhizaria with Chromalveolates. Mol Biol Evol. 2007
- McFadden GI. Primary and secondary endosymbiosis and the origin of plastids. J Phycol. 2001
- Rodriguez-Ezpeleta N, Brinkmann H, Burey SC, Roure B, Burger G, et al. Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr Biol. 2005
- Nosenko T, Bhattacharya D. Horizontal gene transfer in chromalveolates. BMC Evol Biol. 2007
- Lane CE, van den Heuvel K, Korera C, Curtis BA, Parsons BJ, et al. Nucleomorph genome of Hemiselmis andersenii reveals complete intron loss and compaction as a driver of protein structure and function. Proc Natl Acad Sci U S A. 2007
- Douglas S, Zauner S, Fraunholz M, Beaton M, Penny S, et al. The highly reduced genome of an enslaved algal nucleus. Nature. 2001
- Vørs N. Ultrastructure and autecology of the marine, heterotrophic flagellate Leucocryptos marina (Braaud) Butcher 1967 (Kathablepharidaceae/Kathablepharidae), with a discussion of the genera Leucocryptos and Katablepharis/Kathablepharis. Eur J Protistol. 1992
- McFadden GI, Gilson PR, Hill DRA. Goniomonas: ribosomal RNA sequences indicate that this phagotrophic flagellate is a close relative of the host component of cryptomonads. Eur J Phycol. 1994
- Maddison WP. Gene trees in species trees. Syst Biol. 1997
- Stiller JW. Plastid endosymbiosis, genome evolution and the origin of green plants. Trends Plant Sci. 2007
- Steiner JM, Yusa F, Pompe JA, Loffelhardt W. Homologous protein import machineries in chloroplasts and cyanelles. Plant J. 2005
- Stoebe B, Kowallik KV. Gene-cluster analysis in chloroplast genomics. Trends Genet. 1999
- Durnford DG, Deane JA, Tan S, McFadden GI, Gantt E, et al. A phylogenetic assessment of the eukaryotic light-harvesting antenna proteins, with implications for plastid evolution. J Mol Evol. 1999
- Rissler HM, Durnford DG. Isolation of a novel carotenoid-rich protein in Cyanophora paradoxa that is immunologically related to the light-harvesting complexes of photosynthetic eukaryotes. Plant Cell Physiol. 2005
- Stoebe B, Martin W, Kowallik KV. Distribution and nomenclature of protein-coding genes in 12 sequenced chloroplast genomes. Plant Mol Biol Rep. 1998
- Loffelhardt W, Bohnert HJ, Bryant DA. The complete sequence of the Cyanophora paradoxa cyanelle genome (Glaucocystophyceae). Plant Syst Evol. 1997
- O"Kelly C. Relationships of eukaryotic algal groups to other protists. In: Berner T, editor. Ultrastructure of microalgae. Boca Raton, FL: CRC Press; 1993
- Stiller JW, Harrell L. The largest subunit of RNA polymerase II from the Glaucocystophyta: functional constraint and short-branch exclusion in deep eukaryotic phylogeny. BMC Evol Biol. 2005
- Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science. 2000
- Burger G, Saint-Louis D, Gray MW, Lang BF. Complete sequence of the mitochondrial DNA of the red alga Porphyra purpurea: cyanobacterial introns and shared ancestry of red and green algae. Plant Cell. 1999
- Secq MPO, Goer SL, Stam WT, Olsen JL. Complete mitochondrial genomes of the three brown algae (Heterokonta: Phaeophyceae) Dictyota dichotoma, Fucus vesiculosus and Desmarestia viridis. Curr Genet. 2006
- Kim E, Lane CE, Curtis BA, Kozera C, Bowman S, et al. Complete sequence and analysis of the mitochondrial genome of Hemiselmis andersenii CCMP644 (Cryptophyceae). BMC Genomics. 2008
- Gibbs SP. The Chloroplasts of some algal groups may have evolved from endosymbiotic eukaryotic algae. Ann N Y Acad Sci. 1981
- Rumpho ME, Summer EJ, Manhart JR. Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis. Plant Physiol. 2000
- Leander BS, Keeling PJ. Morphostasis in alveolate evolution. Trends Ecol Evol. 2003
- Moriya M, Nakayama T, Inouye I. A new class of the stramenopiles, Placididea classis nova: description of Placidia cafeteriopsis gen. et sp nov. Protist. 2002
- Kim E, Archibald JM. Diversity and evolution of plastids and their genomes. In: Sandelius AS, Aronsson H, editors. The Chloroplast: Interactions with the environment. Heidelberg: Springer; 2008
- Harper JT, Keeling PJ. Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol Biol Evol. 2003
- Takishita K, Ishida KI, Maruyama T. Phylogeny of nuclear-encoded plastid-targeted GAPDH gene supports separate origins for the peridinin- and the fucoxanthin derivative-containing plastids of dinoflagellates. Protist. 2004
- Takishita K, Kawachi M, Noel MH, Matsumoto T, Kakizoe N, et al. Origins of plastids and glyceraldehyde-3-phosphate dehydrogenase genes in the green-colored dinoflagellate Lepidodinium chlorophorum. Gene. 2008
- Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, et al. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc Natl Acad Sci U S A. 2002
- Ohta N, Matsuzaki M, Misumi O, Miyagishima S, Nozaki H, et al. Complete sequence and analysis of the plastid genome of the unicellular red alga Cyanidioschyzon merolae. DNA Res. 2003
- Bachvaroff TR, Puerta MVS, Delwiche CF. Chlorophyll c-containing plastid relationships based on analyses of a multigene data set with all four chromalveolate lineages. Mol Biol Evol. 2005
- Bodyl A, Moszczynski K. Did the peridinin plastid evolve through tertiary endosymbiosis? A hypothesis. Eur J Phycol. 2006
- Lee RE, Kugrens P. Katablepharis ovalis, a colorless flagellate with interesting cytological characteristics. J Phycol. 1991
- Lee RE, Kugrens P, Mylnikov AP. The structure of the flagellar apparatus of two strains of Katablepharis (Cryptophyceae). Br Phycol J. 1992
- Clay B, Kugrens P. Systematics of the enigmatic kathablepharids, including EM characterization of the type species, Kathablepharis phoenikoston, and new observations on K. remigera com. nov. Protist. 1999
- Domozych DS, Wells B, Shaw PJ. Scale biogenesis in the green alga, Mesostigma viride. Protoplasma. 1992
- Domozych DS, Stewart KD, Mattox KR. Development of the cell wall in Tetraselmis: role of the Golgi apparatus and extracellular wall assembly. J Cell Sci. 1981
- Gupta RS. Protein phylogenies and signature sequences: a reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiol Mol Biol Rev. 1998
- Boorstein WR, Ziegelhoffer T, Craig EA. Molecular evolution of the HSP70 multigene family. J Mol Evol. 1994
- Maddison DR, Maddison WP. MacClade 4: analysis of phylogeny and character evolution. Sunderland, MA: Sinauer Associates Inc; 2001
- Inagaki Y, Simpson AGB, Dacks JB, Roger AJ. Phylogenetic artifacts can be caused by leucine, serine, and arginine codon usage heterogeneity: dinoflagellate plastid origins as a case study. Syst Biol. 2004
- Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006
- Lartillot N, Brinkmann H, Philippe H. Suppression of long-branch attraction artefacts in the animal phylogeny using a site-heterogeneous model. BMC Evol Biol. 2007
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005
- Schmidt HA, Strimmer K, Vingron M, von Haeseler A. TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics. 2002
- Desper R, Gascuel O. Fast and accurate phylogeny reconstruction algorithms based on the minimum-evolution principle. J Comput Biol. 2002
- Felsenstein J. Seattle: Department of Genome Sciences, University of Washington; 2005
Литература на русском
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, том 47, вып. 2, с. 103-120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова . - М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341-356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3-12.
- Б. М. Медников. Биология: формы и уровни жизни. - Просвещение, 1995.
- Д. Тейлор, Н. Грин, У. Стаут. Биология (в трёх томах)
- В.В.Малахов. Основные этапы эволюции эукариотных организмов. 2003
- М. А. Федонкин. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь. 2003
- С. В. Шестаков. О ранних этапах биологической эволюции с позиции геномики. 2003
- Марков А.В. Проблема происхождения эукариот
- А.В.Марков, А.М.Куликов. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы
- Г.А.Заварзин. Эволюция микробных сообществ.
- Н.А.Колчанов. Эволюция регуляторных генетических систем.
- А.Ю.Розанов, М.А.Федонкин. Проблема первичного биотопа эвкариот. 1994.
- Ю.Ф.Богданов, С.Я.Дадашев, Т.М.Гришаева. Сравнительная геномика и протеомика дрозофилы, нематоды Бреннера и арабидопсиса. Идентификация функционально сходных генов и белков синапсиса мейотических хромосом
- Ермилова Е.В., Залуцкая Ж.М., Лапина Т.В. Подвижность и поведение микроорганизмов Т.2: Эукариоты
- Греннер Д., Марри Р., Мейес П., Родуэлл В. Биохимия человека
Arabic Bulgarian Chinese Croatian Czech Danish Dutch English Estonian Finnish French German Greek Hebrew Hindi Hungarian Icelandic Indonesian Italian Japanese Korean Latvian Lithuanian Malagasy Norwegian Persian Polish Portuguese Romanian Russian Serbian Slovak Slovenian Spanish Swedish Thai Turkish Vietnamese
Эукариоты
Материал из Википедии - свободной энциклопедии
| Ядерные |
|---|
| Научная классификация |
| Классификация: Организмы Надцарство: Эукариоты |
| Латинское название |
| Eucaryota |
| Царства |
| На Викивидах есть страница по этой теме |
| На Викискладе есть страница по этой теме |
Диаграмма типичной клетки животного. Отмеченные органоиды (органеллы) 1. Ядрышко 2. Ядро 3. Рибосома 4. Везикула 5. Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат Гольджи 7. Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический ретикулум 9. Митохондрия 10. Вакуоль 11. Гиалоплазма 12. Лизосома 13. Центросома (Центриоль)
Эукарио́ты , или Я́дерные (лат. Eucaryota от греч. εύ- - хорошо и κάρυον - ядро) - надцарство живых организмов , клетки которых содержат ядра . Все организмы, кроме бактерий и археев , являются ядерными.
Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических , разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы , что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана , рибосомы (существенно отличные от эукариотических рибосом) и генетический материал - бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро - это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК , «упакованные» в хромосомы . Ядро обычно одно, но бывают и многоядерные клетки.
Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных . Затем было выделено царство грибов , которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших , миксомицетов , хромистов . Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона - Unikonta и Bikonta .
Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески "эукариот" значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки - нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды . По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза , в том числе у многих групп - фагоцитоза . Фагоцитозом (дословно "поедание клеткой") называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И.И. Мечниковым у морских звезд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники .
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты , грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов , диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина . Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию, для которых отпала.
Ещё одно отличие - строение жгутиков. У бактерий они тонкие - всего 15-20 нм в диаметре. Это полые нити из белка флагеллина . Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5-10 мкм, когда тот же показатель у эукариот составляет 10-100 мкм. Объём такой клетки в 1000-10000 раз больше, чем прокариотической. У прокариот рибосомы мелкие (70S-типа). У эукариот рибосомы более крупные (80S-типа).