Specificitatea structurii primare a proteinei. Caracteristicile formării legăturilor peptidice. Rolul determinant al structurii primare în formarea nivelurilor superioare de organizare a moleculei proteice.
Structura primară a unei proteine.
Structura primară a proteinelor este un lanț polipeptidic liniar de aminoacizi conectați prin legături peptidice. Structura primară este cel mai simplu nivel de organizare structurală a unei molecule de proteine. Stabilitate ridicată îi este dată de legăturile peptidice covalente dintre gruparea α-amino a unui aminoacid și gruparea α-carboxil a unui alt aminoacid.
Dacă gruparea imino a prolinei sau hidroxiprolinei este implicată în formarea unei legături peptidice, atunci are o formă diferită.
Când se formează legături peptidice în celule, gruparea carboxil a unui aminoacid este mai întâi activată și apoi se combină cu gruparea amino a altuia. Sinteza de laborator a polipeptidelor se realizează aproximativ în același mod.
O legătură peptidică este un fragment repetat al unui lanț polipeptidic. Are o serie de caracteristici care afectează nu numai forma structurii primare, ci și nivelurile superioare de organizare a lanțului polipeptidic:
Coplanaritate - toți atomii incluși în grupul peptidic sunt în același plan;
Capacitatea de a exista în două forme de rezonanță (forma ceto sau enol);
Poziția trans a substituenților față de legătura C-N;
Capacitatea de a forma legături de hidrogen și fiecare dintre grupele peptidice poate forma două legături de hidrogen cu alte grupări, inclusiv cu cele peptidice.
Excepție fac grupele peptidice care implică gruparea amino a prolinei sau hidroxiprolinei. Ei sunt capabili să formeze o singură legătură de hidrogen (vezi mai sus). Acest lucru afectează formarea structurii secundare a proteinei. Lanțul polipeptidic din zona în care se află prolina sau hidroxiprolina se îndoaie ușor, deoarece nu este ținut, ca de obicei, de o a doua legătură de hidrogen.
4. Structura secundară a proteinelor. Legături care stabilizează structura secundară, α-helix. Factorii care perturbă spiralarea. Structura foii β, caracteristicile structurii conformaționale.
Structura secundară a unei proteine este configurația lanțului polipeptidic, adică. o metodă de pliere a unui lanț polipeptidic într-o conformație specifică.
Acest proces nu are loc aleatoriu, ci în conformitate cu structura primară a proteinei.
Structura secundară este menținută în primul rând prin legături de hidrogen, deși pentru unele proteine legăturile covalente peptidice și disulfurice au o anumită contribuție.
Cel mai probabil tip de structură secundară a proteinelor globulare este α-helix. Lanțul polipeptidic se răsucește într-o spirală în sensul acelor de ceasornic. Fiecare proteină este caracterizată printr-un anumit grad de elicoidalizare. Astfel, lanțurile polipeptidice ale hemoglobinei sunt spiralate cu 75%, iar molecula de pepsină cu 30%.
Tipul de configurație a lanțurilor polipeptidice atunci când segmentele lanțului peptidic sunt aranjate într-un singur strat, formând o structură similară unei foi pliate într-un acordeon, se numește structură . Acest tip de structură secundară se găsește în proteinele din mușchi, păr și mătase. Stratul poate fi intramolecular sau format din două sau mai multe lanțuri polipeptidice.
Capacitatea de a forma legături de hidrogen, care sunt forța motrice pentru formarea structurilor α și β într-o moleculă de proteină, este exprimată în grade diferite în diferiți aminoacizi. Există un grup de aminoacizi care formează spirală: ala, gln, glu, leu, met, lys, gis. Val, ile, tyr, tre, phen contribuie la formarea structurilor ale lanțului polipeptidic. Prezența sulfului, gly, pro, asn, asp duce la formarea predominantă a fragmentelor dezordonate în molecula proteică.
Există proteine în natură a căror structură nu corespunde cu niciuna
β-, nici -structură (colagen).
5.Structura terțiară a proteinei. Legături care stabilizează structura terțiară (covalentă, ionică, hidrofobă, hidrogen, van der Waals).
Structura terțiară a unei proteine este orientarea spațială a helixului polipeptidic sau modul în care lanțul polipeptidic este așezat într-un anumit volum. Prima proteină, structura terțiară a proteinei (mioglobina de cașlot) a fost stabilită pentru prima dată prin analiza de difracție cu raze X (Fig. 2).
În stabilizarea structurii spațiale a proteinelor, pe lângă legăturile covalente, rolul principal revine legăturilor necovalente (forțe intermoleculare van der Waals, legături de hidrogen, interacțiuni electrostatice ale grupărilor ionizate, interacțiuni hidrofobe etc.).
Cu ajutorul analizei de difracție cu raze X s-a stabilit existența unor niveluri specifice de organizare structurală a moleculei proteice, intermediare între structurile secundare și terțiare. Un domeniu este o unitate structurală globulară compactă în cadrul unui lanț polipeptidic (Fig. 3). Au fost descoperite proteine (în special imunoglobuline) în care există domenii cu structură și funcții diferite.
Conform conceptelor moderne, după terminarea sintezei proteinelor, structura sa terțiară se formează spontan. Procesul de formare a structurii spațiale native a unui lanț polipeptidic este pliere. Principala forță motrice a plierii este interacțiunea radicalilor de aminoacizi cu moleculele de apă. În acest caz, radicalii hidrofobi de aminoacizi sunt orientați în interiorul moleculei proteice, iar radicalii hidrofili sunt îndreptați către apă.
Celulele conțin proteine numite chaperone. Funcția lor principală este participarea la pliere (Fig. 4). Au fost descrise o serie de boli umane de natură ereditară, a căror apariție este asociată cu perturbarea procesului de pliere din cauza mutațiilor (pigmentoză, fibroză etc.).
Toate proprietățile biologice ale proteinelor sunt asociate cu formarea și conservarea unei structuri terțiare numită nativă. Globulul proteic nu este o structură absolut rigidă: sunt posibile mișcări reversibile ale fragmentelor din lanțul polipeptidic. Aceste modificări nu duc la perturbarea conformației generale a moleculei. Factorii care influențează conformația unei molecule proteice sunt tăria ionică a soluției, pH-ul mediului, interacțiunea cu componentele soluției. Orice influență care duce la perturbarea structurii native a moleculei duce la pierderea parțială sau completă a proprietăților biologice ale proteinei.
6. Structura cuaternară a proteinei. Conceptul de monomeri și oligomeri. Dependența proprietăților proteinei de conformația sa. Relația dintre structură și funcție.
Structura cuaternară a unei proteine este aranjarea lanțurilor polipeptidice individuale cu o structură specifică primară, secundară sau terțiară în spațiu și formarea unei singure formațiuni macromoleculare.
O proteină formată din mai multe lanțuri polipeptidice se numește oligomer, iar fiecare lanț polipeptidic inclus în ea se numește protomer. Proteinele oligomerice, de regulă, constau dintr-un număr par de subunități, de exemplu, molecula de hemoglobină este construită din două lanțuri α- și două β-polipeptide (Fig. 5).
Aproximativ 5% dintre proteine, cum ar fi feritina și imunoglobulinele, au o structură cuaternară. Structura subunității este caracteristică multor enzime, în primul rând celor care îndeplinesc funcții complexe. Aproape toate polimerazele ADN și ARN au o structură cuaternară. Lanțurile polipeptidice care alcătuiesc o proteină cu structură cuaternară se formează separat pe ribozomi. Numai după terminarea sintezei se combină într-o structură supramoleculară. Proteina capătă activitate biologică la nivelul structurii cuaternare. Stabilizarea structurii cuaternare are loc cu participarea acelorași legături ca și în timpul formării structurii terțiare, cu excepția legăturilor covalente.
O serie de cercetători recunosc existența unui al cincilea nivel de organizare structurală a proteinelor. Complexele macromoleculare polifuncționale ale diferitelor enzime care catalizează întreaga cale a transformărilor substratului se numesc metaboloni (complex de piruvat dehidrogenază, sintetaze IVH, lanț respirator).
O proteină care îndeplinește o funcție specifică în metabolismul celular poate fi reprezentată sub mai multe forme - proteine izofuncționale, sau izoproteine. Mai multe forme de hemoglobină se găsesc în globulele roșii umane: La un adult, forma predominantă este HbA. Ch Hemoglobina fetală HbF este caracteristică stadiului embrionar al dezvoltării umane. Toate formele de hemoglobine îndeplinesc funcția de a transporta oxigenul de la plămâni la țesuturi, dar proprietățile diferitelor hemoglobine diferă.
Conceptul de proteină nativă. Conceptul de proteine alosterice.
Proteina nativă este o proteină care are o anumită activitate biologică.
8. Principalele funcții ale proteinelor simple și complexe din organism: structurale, catalitice, receptor, de reglare, de transport, de protecție, contractile și altele.
Funcția structurală. În combinație cu lipidele, proteinele participă la formarea biomembranelor celulare. Proteinele structurale ale citoscheletului dau formă celulelor și multor organite. Exemple de proteine structurale sunt colagenul din țesutul conjunctiv, keratina din păr, piele și unghii.
Funcția contractilă (motorie). Funcția contractilă este îndeplinită de proteinele musculare (actina și miozina). Proteinele citoscheletice sunt necesare pentru segregarea cromozomilor în timpul mitozei.
Funcția nutrițională (rezervă). Ovalbumina (albușurile de ou) sunt surse de nutriție pentru făt. Cazeina, o proteină din lapte, are și o funcție nutritivă.
Funcția catalitică. Cele mai multe enzime cunoscute în prezent (catalizatori biologici) sunt proteine.
Funcția de transport. Hemoglobina proteică din hematii este implicată în transportul oxigenului și dioxidului de carbon, îndeplinind funcția respiratorie. Albuminele serice sunt implicate în transportul lipidelor.
Funcție de protecție. Ca răspuns la intrarea în organism a virușilor, bacteriilor, proteinelor străine și toxinelor, se formează proteine de protecție - anticorpi (apărare imunitară). Proteinele plasmatice specifice sunt capabile de coagulare, ceea ce protejează împotriva pierderii de sânge în timpul sângerării (protecție fizică).
Funcția receptorului. Proteinele celulare formează receptori specifici și participă la transmiterea semnalelor hormonale.
Funcția hormonală. Un grup de hormoni sunt proteine sau polipeptide, de exemplu, hormonul hipofizar vasopresina.
Alte funcții importante ale proteinelor sunt proprietățile de tamponare (asigurarea pH-ului fiziologic al mediului intern), menținerea presiunii oncotice în celule și sânge etc.
În corpul oamenilor și al animalelor, conținutul de proteine este mult mai mare decât la plante. În mușchi, plămâni, splină, rinichi, proteinele reprezintă mai mult de 70-80% din masa uscată în ficat - 57%, în creier - 45%. Conținut scăzut de proteine în oase și dinți - 20 și 18%. Conținut inegal de proteine în diferite organite subcelulare. Există mai multe proteine în hialoplasmă (sucul intracelular). Dacă luăm proteina totală din celule ca fiind 100%, atunci hialoplasma reprezintă 40%. Mitocondriile și microzomii conțin fiecare 20%, nucleul - 12%, lizozomii - 2%, peroxizomii - 2,5%, membrana plasmatică - 1,5% proteine.
Unele proteine conțin fosfor (0,2-2%), fier și alte elemente. Cel mai constant conținut de azot pentru proteinele de origine animală, vegetală și microbiană este în medie de 16%; pe această bază, cantitatea de proteină este calculată din conținutul de azot: masa de azot determinată de analiză este înmulțită cu un factor de 6,25 (100:16 = 6,25).
Dimensiunea moleculelor de proteine variază de la 1 micron la 1 nm
Metoda de filtrare pe gel sau sită moleculară
Pentru separarea proteinelor se folosesc adesea metode cromatografice, bazate pe distribuția substanțelor între două faze, dintre care una mobilă și cealaltă imobilă. Metodele cromatografice se bazează pe diferite principii: filtrare pe gel, schimb ionic, adsorbție, afinitate biologică.
Metoda de separare a proteinelor folosind cromatografia de filtrare pe gel se bazează pe faptul că substanțele care diferă ca greutate moleculară sunt distribuite diferit între faza staționară și cea mobilă. Coloana cromatografică este umplută cu granule dintr-o substanță poroasă (Sephadex, agaroză etc.). În structura polizaharidei, se formează legături încrucișate și se formează granule cu „pori”, prin care trece cu ușurință apa și substanțele cu molecularitate scăzută. În funcție de condiții, se pot forma granule cu dimensiuni diferite de „pori”.
Faza staționară este lichidul din interiorul granulelor, în care pot pătrunde substanțele cu greutate moleculară mică și proteinele cu greutate moleculară mică. Amestecul de proteine aplicat pe coloana de cromatografie este spălat (eluat) prin trecerea unui solvent prin coloană. Cele mai mari molecule se mișcă, de asemenea, împreună cu frontul de solvent.
Moleculele mai mici difuzează în granulele Sephadex și intră în faza staționară pentru o perioadă de timp, drept urmare mișcarea lor este întârziată. Mărimea porilor determină mărimea moleculelor care pot pătrunde în interiorul granulelor (Fig. 1-55).
Deoarece structura gelului Sephadex este ușor deformată sub presiune, gelurile au început să fie înlocuite cu matrici mai rigide (Sefactil, Toy-Operl), care sunt granule sferice cu dimensiuni diferite ale porilor. Alegerea dimensiunilor porilor în granule depinde de scopurile cromatografiei (alte metode cromatografice vor fi discutate mai jos).
Proteine native și non-native
Proteinele native sunt cele care conțin toți aminoacizii esențiali de care organismul are nevoie pentru a construi și repara mușchii și organele.
Proteinele non-native sunt cele care conțin doar o parte din aminoacizi, dar au totuși o valoare nutritivă semnificativă.
Proteinele native se găsesc în carne, pește, fructe de mare, păsări de curte, ouă și brânză. De asemenea, sunt bogate în vitamine B.
Proteinele non-native se găsesc în cereale, leguminoase, nuci, semințe și unele legume cu frunze. Și, de asemenea, în unturi de nuci precum arahide, migdale și caju.
Proteinele non-native sunt utile pentru a fi consumate în combinație cu alte alimente. Mâncând combinații de anumite proteine non-native, puteți obține toți aminoacizii esențiali într-o singură masă.
Din cartea Ortotrofie: elementele de bază ale nutriției adecvate și postului terapeutic autor Herbert McGolfin Shelton Din cartea Codul femeii de Alice Vitti Din cartea Nutriție și dietă pentru sportivi autor Elena Anatolievna Boyko Din cartea Stretching for Health and Longevity autor Vanessa Thompson Din cartea Rețete reale împotriva celulitei.5 minute pe zi autor Kristina Aleksandrovna Kulagina Din cartea Diabet. Prevenirea, diagnosticarea și tratamentul folosind metode tradiționale și netradiționale autor Violetta Romanovna Khamidova Din cartea The Hollywood Diet autor D. B. Abramov Din cartea Cum să nu te transformi în Baba Yaga de doctorul Nonna Din cartea Pocket Calorie Counter autor Iulia Luzhkovskaya Din cartea Obiceiuri sănătoase. Dieta doctorului Ionova autoarea Lydia IonovaMODULUL 1 STRUCTURA, PROPRIETĂȚI ȘI FUNCȚII ALE PROTEINELOR
MODULUL 1 STRUCTURA, PROPRIETĂȚI ȘI FUNCȚII ALE PROTEINELOR
Structura modulului | Teme |
Unitate modulară 1 | 1.1. Organizarea structurală a proteinelor. Etape de formare a conformației native a proteinelor 1.2. Bazele funcționării proteinelor. Medicamentele ca liganzi care afectează funcția proteinelor 1.3. Denaturarea proteinelor și posibilitatea renativării lor spontane |
Unitatea modulară 2 | 1.4. Caracteristici ale structurii și funcționării proteinelor oligomerice folosind exemplul hemoglobinei 1.5. Menținerea conformației proteinelor native în condiții celulare 1.6. Varietate de proteine. Familii de proteine folosind exemplul imunoglobulinelor 1.7. Proprietățile fizico-chimice ale proteinelor și metodele de separare a acestora |
Unitatea modulara 1 ORGANIZAREA STRUCTURALA A PROTEINELOR MONOMERICE SI BAZELE FUNCTIONARII LOR
Obiective de învățare A fi capabil să:
1. Utilizați cunoștințele despre caracteristicile structurale ale proteinelor și dependența funcțiilor proteinelor de structura lor pentru a înțelege mecanismele de dezvoltare a proteinopatiilor ereditare și dobândite.
2. Explicați mecanismele acțiunii terapeutice a unor medicamente ca liganzi care interacționează cu proteinele și își modifică activitatea.
3. Utilizați cunoștințele despre structura și labilitatea conformațională a proteinelor pentru a înțelege instabilitatea lor structurală și funcțională și tendința de denaturare în condiții în schimbare.
4. Explicați utilizarea agenților de denaturare ca mijloace de sterilizare a materialelor și instrumentelor medicale, precum și a antisepticelor.
Știi:
1. Niveluri de organizare structurală a proteinelor.
2. Importanța structurii primare a proteinelor, care determină diversitatea lor structurală și funcțională.
3. Mecanismul de formare a centrului activ în proteine și interacțiunea specifică a acestuia cu ligand, care stă la baza funcționării proteinelor.
4. Exemple de influență a liganzilor exogeni (medicamente, toxine, otrăvuri) asupra conformației și activității funcționale a proteinelor.
5. Cauze si consecinte ale denaturarii proteinelor, factori care determina denaturarea.
6. Exemple de utilizare a factorilor denaturanți în medicină ca antiseptice și mijloace pentru sterilizarea instrumentelor medicale.
TEMA 1.1. ORGANIZAREA STRUCTURALĂ A PROTEINELOR. ETAPE DE FORMARE A NATIVULUI
CONFORMATII PROTEINE
Proteinele sunt molecule polimerice ai căror monomeri sunt doar 20 de α-aminoacizi. Setul și ordinea combinației de aminoacizi într-o proteină este determinată de structura genelor din ADN-ul indivizilor. Fiecare proteină, în conformitate cu structura sa specifică, își îndeplinește propria funcție. Setul de proteine ale unui anumit organism determină caracteristicile sale fenotipice, precum și prezența bolilor ereditare sau o predispoziție la dezvoltarea lor.
1. Aminoacizi care alcătuiesc proteinele. Legătură peptidică. Proteinele sunt polimeri formați din monomeri - 20 de α-aminoacizi, a căror formulă generală este
Aminoacizii diferă în ceea ce privește structura, dimensiunea și proprietățile fizico-chimice ale radicalilor atașați la atomul de carbon α. Grupurile funcționale ale aminoacizilor determină caracteristicile proprietăților diferiților α-aminoacizi. Radicalii găsiți în α-aminoacizi pot fi împărțiți în mai multe grupuri:
prolina, Spre deosebire de ceilalți 19 monomeri proteici, nu este un aminoacid, ci un iminoacid; radicalul din prolină este asociat atât cu atomul de carbon α, cât și cu gruparea imino.
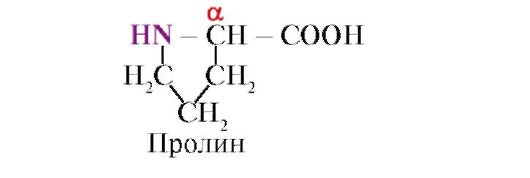 Aminoacizii variază în solubilitate în apă. Acest lucru se datorează capacității radicalilor de a interacționa cu apa (hidrat).
Aminoacizii variază în solubilitate în apă. Acest lucru se datorează capacității radicalilor de a interacționa cu apa (hidrat).
LA hidrofil includ radicali care conțin grupări funcționale neîncărcate anionice, cationice și polare.
LA hidrofob includ radicali care conțin grupări metil, lanțuri alifatice sau inele.
2. Legăturile peptidice conectează aminoacizii pentru a forma peptide.În timpul sintezei peptidelor, gruparea α-carboxil a unui aminoacid interacționează cu gruparea α-amino a altui aminoacid pentru a forma legătură peptidică:
Proteinele sunt polipeptide, adică. polimeri liniari ai α-aminoacizilor legați printr-o legătură peptidică (Fig. 1.1.)
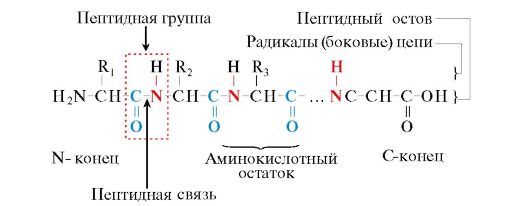 Orez. 1.1. Termeni folosiți pentru a descrie structura peptidelor
Orez. 1.1. Termeni folosiți pentru a descrie structura peptidelor
Monomerii aminoacizilor care formează polipeptidele se numesc reziduuri de aminoacizi. Un lanț de grupuri care se repetă - NH-CH-CO- forme coloana vertebrală peptidică. Un rest de aminoacid care are o grupare α-amino liberă se numește N-terminal, iar unul care are o grupare α-carboxil liberă se numește C-terminal. Peptidele sunt scrise și citite de la capătul N-terminal la capătul C-terminal.
Legătura peptidică formată de grupul imino a prolinei diferă de alte legături peptidice: atomul de azot al grupului peptidic nu are hidrogen,
în schimb, există o legătură cu un radical, ca urmare a căreia o parte a inelului este inclusă în scheletul peptidic:
 Peptidele diferă în ceea ce privește compoziția de aminoacizi, numărul de aminoacizi și ordinea conexiunii aminoacizilor, de exemplu, Ser-Ala-Glu-Gis și His-Glu-Ala-Ser sunt două peptide diferite.
Peptidele diferă în ceea ce privește compoziția de aminoacizi, numărul de aminoacizi și ordinea conexiunii aminoacizilor, de exemplu, Ser-Ala-Glu-Gis și His-Glu-Ala-Ser sunt două peptide diferite.
Legăturile peptidice sunt foarte puternice, iar hidroliza lor chimică neenzimatică necesită condiții dure: proteina analizată este hidrolizată în acid clorhidric concentrat la o temperatură de aproximativ 110° timp de 24 de ore. Într-o celulă vie, legăturile peptidice pot fi rupte enzime proteolitice, numit proteaze sau hidrolaze peptidice.
3. Structura primară a proteinelor. Reziduurile de aminoacizi din lanțurile peptidice ale diferitelor proteine nu alternează aleatoriu, ci sunt aranjate într-o anumită ordine. Se numește secvența liniară sau ordinea de alternanță a resturilor de aminoacizi dintr-un lanț polipeptidic structura primară a proteinei.
Structura primară a fiecărei proteine individuale este codificată în molecula de ADN (într-o regiune numită genă) și este realizată în timpul transcripției (copierea informațiilor pe ARNm) și translației (sinteza structurii primare a proteinei). În consecință, structura primară a proteinelor unei persoane individuale este informația transmisă ereditar de la părinți la copii, care determină caracteristicile structurale ale proteinelor unui anumit organism, de care depinde funcția proteinelor existente (Fig. 1.2.).
 Orez. 1.2. Relația dintre genotip și conformația proteinelor sintetizate în corpul individului
Orez. 1.2. Relația dintre genotip și conformația proteinelor sintetizate în corpul individului
Fiecare dintre cele aproximativ 100.000 de proteine individuale din corpul uman are unic structura primara. Moleculele de același tip de proteină (de exemplu, albumina) au aceeași alternanță de resturi de aminoacizi, ceea ce distinge albumina de orice altă proteină individuală.
Secvența de resturi de aminoacizi dintr-un lanț peptidic poate fi considerată o formă de înregistrare a informațiilor. Aceste informații determină aranjarea spațială a lanțului peptidic linear într-o structură tridimensională mai compactă numită conformaţie veveriţă. Procesul de formare a unei conformații proteice active funcțional se numește pliere
4. Conformația proteinei. Rotația liberă în scheletul peptidic este posibilă între atomul de azot al grupării peptidice și atomul de carbon α vecin, precum și între atomul de carbon α și carbonul grupării carbonil. Datorită interacțiunii grupurilor funcționale de reziduuri de aminoacizi, structura primară a proteinelor poate dobândi structuri spațiale mai complexe. În proteinele globulare, există două niveluri principale de pliere a conformației lanțurilor peptidice: secundarȘi structura tertiara.
Structura secundară a proteinelor este o structură spațială formată ca urmare a formării legăturilor de hidrogen între grupările funcționale -C=O și -NH- ale scheletului peptidic. În acest caz, lanțul peptidic poate dobândi structuri regulate de două tipuri: α-helicesȘi β-structuri.
ÎN α-helices se formează legături de hidrogen între atomul de oxigen al grupării carbonil și hidrogenul azotului amidic al celui de-al 4-lea aminoacid din acesta; lanțurile laterale de reziduuri de aminoacizi
sunt situate de-a lungul periferiei spiralei, fără a participa la formarea structurii secundare (Fig. 1.3.).
Radicalii în vrac, sau radicalii care poartă sarcini egale, împiedică formarea unui α-helix. Reziduul de prolină, care are o structură de inel, întrerupe α-helix, deoarece din cauza lipsei de hidrogen la atomul de azot din lanțul peptidic este imposibil să se formeze o legătură de hidrogen. Legătura dintre azot și atomul de carbon α face parte din inelul de prolină, astfel încât coloana vertebrală a peptidei devine îndoită în acest punct.
β-Structură se formează între regiunile liniare ale scheletului peptidic al unui lanț polipeptidic, formând astfel structuri pliate. Se pot forma lanțuri polipeptidice sau părți ale acestora paralel sau β-structuri antiparalele.În primul caz, terminalele N- și C-terminale ale lanțurilor peptidice care interacționează coincid, iar în al doilea au direcția opusă (Fig. 1.4).
Orez. 1.3. Structura secundară a proteinei - α-helix
 Orez. 1.4. Structuri β-sheet paralele și antiparalele
Orez. 1.4. Structuri β-sheet paralele și antiparalele
Structurile β sunt indicate prin săgeți largi: A - Structura β antiparalelă. B - Structuri paralele β-sheet
În unele proteine, structurile β pot fi formate datorită formării legăturilor de hidrogen între atomii scheletului peptidic al diferitelor lanțuri polipeptidice.
Se găsește și în proteine zone cu secundar neregulat structura, care include îndoituri, bucle și întoarceri ale coloanei vertebrale polipeptidice. Ele sunt adesea localizate în locuri în care direcția lanțului peptidic se schimbă, de exemplu, atunci când se formează o structură paralelă a foii β.
Pe baza prezenței elicelor α și structurilor β, proteinele globulare pot fi împărțite în patru categorii.
Orez. 1.5. Structura secundară a mioglobinei (A) și a lanțului β a hemoglobinei (B), care conține opt elice α

 Orez. 1.6. Structura secundară a domeniului triozofosfat izomerazei și piruvat kinazei
Orez. 1.6. Structura secundară a domeniului triozofosfat izomerazei și piruvat kinazei
 Orez. 1.7. Structura secundară a domeniului constant al imunoglobulinei (A) și a enzimei superoxid dismutază (B)
Orez. 1.7. Structura secundară a domeniului constant al imunoglobulinei (A) și a enzimei superoxid dismutază (B)
ÎN a patra categorie au inclus proteine care conțin o cantitate mică de structuri secundare regulate. Aceste proteine includ proteine mici sau metaloproteine bogate în cisteină.
Structura terțiară a proteinelor- un tip de conformatie format ca urmare a interactiunilor dintre radicalii de aminoacizi, care pot fi situati la o distanta considerabila unul de altul in lantul peptidic. Majoritatea proteinelor formează o structură spațială asemănătoare unui glob (proteine globulare).
Deoarece radicalii hidrofobi de aminoacizi tind să se combine prin așa-numitele interacțiuni hidrofobeși forțele intermoleculare van der Waals, în interiorul globului proteic se formează un miez hidrofob dens. Radicalii hidrofili ionizați și neionizați sunt localizați în principal pe suprafața proteinei și determină solubilitatea acesteia în apă.
 Orez. 1.8. Tipuri de legături care apar între radicalii de aminoacizi în timpul formării structurii terțiare a unei proteine
Orez. 1.8. Tipuri de legături care apar între radicalii de aminoacizi în timpul formării structurii terțiare a unei proteine
1 - legătură ionică- apare între grupele funcționale încărcate pozitiv și negativ;
2 - legătură de hidrogen- apare intre o grupare hidrofila neincarcata si orice alta grupare hidrofila;
3 - interacțiuni hidrofobe- apar între radicalii hidrofobi;
4 - legătură disulfurică- formată din cauza oxidării grupurilor SH ale reziduurilor de cisteină și a interacțiunii lor între ele
Reziduurile de aminoacizi hidrofile situate în interiorul miezului hidrofob pot interacționa între ele folosind ionicȘi legături de hidrogen(Fig. 1.8).
Legăturile ionice și de hidrogen, precum și interacțiunile hidrofobe, sunt slabe: energia lor nu este cu mult mai mare decât energia mișcării termice a moleculelor la temperatura camerei. Conformația proteinei este menținută prin formarea multor astfel de legături slabe. Deoarece atomii care alcătuiesc o proteină sunt în mișcare constantă, este posibil să se rupă unele legături slabe și să se formeze altele, ceea ce duce la mișcări ușoare ale secțiunilor individuale ale lanțului polipeptidic. Această proprietate a proteinelor de a modifica conformația ca urmare a ruperii unora și a formării altor legături slabe se numește labilitate conformaţională.
Corpul uman are sisteme care susțin homeostaziei- constanța mediului intern în anumite limite acceptabile pentru un organism sănătos. În condiții de homeostazie, mici modificări ale conformației nu perturbă structura și funcția generală a proteinelor. Conformația activă funcțional a unei proteine se numește conformatie nativa. Modificările din mediul intern (de exemplu, concentrația de glucoză, ioni de Ca, protoni etc.) duc la modificări ale conformației și perturbarea funcțiilor proteinelor.
Structura terțiară a unor proteine este stabilizată legături disulfurice, formată ca urmare a interacțiunii grupărilor -SH a două resturi
 Orez. 1.9. Formarea unei legături disulfurice într-o moleculă de proteină
Orez. 1.9. Formarea unei legături disulfurice într-o moleculă de proteină
cisteină (Fig. 1.9). Majoritatea proteinelor intracelulare nu au legături disulfurice covalente în structura lor terțiară. Prezența lor este caracteristică proteinelor secretate de celulă, ceea ce le asigură o mai mare stabilitate în condiții extracelulare. Astfel, legăturile disulfurice sunt prezente în moleculele de insulină și imunoglobuline.
Insulină- un hormon proteic sintetizat în celulele β ale pancreasului și secretat în sânge ca răspuns la creșterea concentrației de glucoză în sânge. În structura insulinei, există două legături disulfurice care conectează lanțurile polipeptidei A și B și o legătură disulfură în cadrul lanțului A (Fig. 1.10).
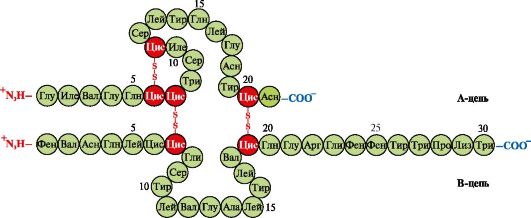 Orez. 1.10. Legături disulfurice în structura insulinei
Orez. 1.10. Legături disulfurice în structura insulinei
5. Structura suprasecundară a proteinelor.În proteinele cu structură și funcții primare diferite, acestea sunt uneori detectate combinații similare și poziții relative ale structurilor secundare, care se numesc structura supersecundara. Ocupă o poziție intermediară între structurile secundare și terțiare, deoarece este o combinație specifică de elemente ale structurii secundare în formarea structurii terțiare a proteinei. Structurile supersecundare au denumiri specifice, cum ar fi „α-helix-turn-a-helix”, „fermoar leucină”, „degete de zinc” etc. Astfel de structuri supersecundare sunt caracteristice proteinelor care leagă ADN-ul.
— Fermoar leucină. Acest tip de structură supersecundară este utilizat pentru a uni două proteine împreună. Pe suprafața proteinelor care interacționează există regiuni elicoidale α care conțin cel puțin patru resturi de leucină. Reziduurile de leucină din α-helix sunt localizate la șase aminoacizi una de cealaltă. Deoarece fiecare rotație a helixului α conține 3,6 reziduuri de aminoacizi, radicalii de leucină sunt localizați pe suprafața fiecărei rotații a doua. Resturile de leucină ale α-helixului unei proteine pot interacționa cu resturile de leucină ale altei proteine (interacțiuni hidrofobe), conectându-le între ele (Fig. 1.11.). Multe proteine care leagă ADN-ul funcționează în complexe oligomerice în care subunitățile individuale sunt legate între ele prin „fermoare cu leucină”.
 Orez. 1.11. „Fermoar leucină” între regiunile α-helicoidale ale două proteine
Orez. 1.11. „Fermoar leucină” între regiunile α-helicoidale ale două proteine
Un exemplu de astfel de proteine sunt histonele. Histones- proteine nucleare, care contin un numar mare de aminoacizi incarcati pozitiv - arginina si lizina (pana la 80%). Moleculele de histonă sunt combinate în complexe oligomerice care conțin opt monomeri folosind „fermoare de leucină”, în ciuda încărcăturii omonime semnificative a acestor molecule.
"Degetul de zinc"- o variantă de structură supersecundară, caracteristică proteinelor de legare a ADN-ului, are forma unui fragment alungit la suprafața proteinei și conține aproximativ 20 de resturi de aminoacizi (Fig. 1.12). Forma „degetului extins” este susținută de un atom de zinc legat de patru radicali de aminoacizi - două reziduuri de cisteină și două reziduuri de histidină. În unele cazuri, în loc de reziduuri de histidină, există reziduuri de cisteină. Două reziduuri de cisteină apropiate sunt separate de celelalte două resturi Gisili printr-o secvență Cys constând din aproximativ 12 resturi de aminoacizi. Această regiune a proteinei formează un α-helix, ai cărui radicali se pot lega în mod specific la regiunile reglatoare ale canelurii majore a ADN-ului. Specificitatea de legare individuală
 Orez. 1.12. Structura primară a regiunii proteinelor care leagă ADN-ul care formează structura „deget de zinc” (literele indică aminoacizii care alcătuiesc această structură)
Orez. 1.12. Structura primară a regiunii proteinelor care leagă ADN-ul care formează structura „deget de zinc” (literele indică aminoacizii care alcătuiesc această structură)
Proteina reglatoare care leagă ADN-ul depinde de secvența reziduurilor de aminoacizi situate în regiunea degetului de zinc. Astfel de structuri conțin, în special, receptori pentru hormonii steroizi implicați în reglarea transcripției (citirea informațiilor de la ADN la ARN).
TEMA 1.2. BAZELE FUNCȚIONĂRII PROTEINELOR. MEDICAMENTE CA LIGANDI AFECTAT FUNCȚIA PROTEINĂ
1. Centrul activ al proteinei și interacțiunea acestuia cu ligand.În timpul formării structurii terțiare, se formează o regiune pe suprafața unei proteine funcțional active, de obicei într-o adâncitură, formată din radicali de aminoacizi care sunt departe unul de celălalt în structura primară. Această regiune, care are o structură unică pentru o anumită proteină și este capabilă să interacționeze în mod specific cu o anumită moleculă sau grup de molecule similare, este numită situs de legare proteină-ligand sau situs activ. Liganzii sunt molecule care interacționează cu proteinele.
Specificitate ridicată Interacțiunea proteinei cu ligand este asigurată de complementaritatea structurii centrului activ cu structura ligandului.
Complementaritatea- aceasta este corespondența spațială și chimică a suprafețelor care interacționează. Centrul activ nu trebuie să corespundă doar spațial ligandului inclus în el, dar și legături (interacțiuni ionice, hidrogen și hidrofobe) trebuie să se formeze între grupările funcționale ale radicalilor incluși în centrul activ și ligand, care dețin ligandul. în centrul activ (Fig. 1.13 ).
 Orez. 1.13. Interacțiunea complementară a proteinei cu ligand
Orez. 1.13. Interacțiunea complementară a proteinei cu ligand
Unii liganzi, atunci când sunt atașați la centrul activ al unei proteine, joacă un rol auxiliar în funcționarea proteinelor. Astfel de liganzi sunt numiți cofactori, iar proteinele care conțin o parte neproteică sunt numite proteine complexe(spre deosebire de proteinele simple, constând numai din partea proteică). Partea non-proteică, ferm conectată la proteină, se numește grupare prostetică. De exemplu, mioglobina, hemoglobina și citocromii conțin un grup protetic, hem, care conține un ion de fier, ferm atașat de centrul activ. Proteinele complexe care conțin hem se numesc hemoproteine.
Atunci când liganzii specifici sunt atașați de proteine, se manifestă funcția acestor proteine. Astfel, albumina, cea mai importantă proteină din plasma sanguină, își manifestă funcția de transport prin atașarea liganzilor hidrofobi, precum acizii grași, bilirubina, unele medicamente etc., la centrul activ (Fig. 1.14)
Liganzii care interacționează cu structura tridimensională a lanțului peptidic pot fi nu numai molecule organice și anorganice cu o moleculă scăzută, ci și macromolecule:
ADN (exemple cu proteine de legare la ADN discutate mai sus);
Polizaharide;
 Orez. 1.14. Relația dintre genotip și fenotip
Orez. 1.14. Relația dintre genotip și fenotip
Structura primară unică a proteinelor umane, codificată în molecula de ADN, este realizată în celule sub forma unei conformații unice, a structurii centrului activ și a funcțiilor proteice.
În aceste cazuri, proteina recunoaște o regiune specifică a ligandului care este proporțională și complementară cu situsul de legare. Astfel, pe suprafața hepatocitelor există proteine receptor pentru hormonul insulină, care are și o structură proteică. Interacțiunea insulinei cu receptorul determină o modificare a conformației sale și activarea sistemelor de semnalizare, ducând la stocarea nutrienților în hepatocite după mese.
Prin urmare, Funcționarea proteinelor se bazează pe interacțiunea specifică a centrului activ al proteinei cu ligand.
2. Structura domeniului și rolul său în funcționarea proteinelor. Lanțurile polipeptidice lungi de proteine globulare se pliază adesea în mai multe regiuni compacte, relativ independente. Au o structură terțiară independentă, care amintește de cea a proteinelor globulare și sunt numite domenii. Datorită structurii de domeniu a proteinelor, structura lor terțiară este mai ușor de format.
În proteinele de domeniu, situsurile de legare a ligandului sunt adesea localizate între domenii. Astfel, tripsina este o enzimă proteolitică care este produsă de partea exocrină a pancreasului și este necesară pentru digestia proteinelor alimentare. Are o structură cu două domenii, iar centrul de legare a tripsinei cu ligandul său - proteina alimentară - este situat în șanțul dintre cele două domenii. În centrul activ, sunt create condițiile necesare pentru legarea eficientă a unui anumit situs al proteinei alimentare și hidroliza legăturilor sale peptidice.
Diferitele domenii dintr-o proteină se pot deplasa unul față de celălalt atunci când centrul activ interacționează cu ligand (Fig. 1.15).
Hexokinaza- o enzima care catalizeaza fosforilarea glucozei folosind ATP. Locul activ al enzimei este situat în despicatură dintre cele două domenii. Când hexokinaza se leagă de glucoză, domeniile din jurul acesteia se închid și substratul devine prins, unde are loc fosforilarea (vezi Fig. 1.15).
 Orez. 1.15. Legarea domeniilor hexokinazei la glucoză
Orez. 1.15. Legarea domeniilor hexokinazei la glucoză
În unele proteine, domeniile îndeplinesc funcții independente prin legarea la diferiți liganzi. Astfel de proteine sunt numite multifuncționale.
3. Medicamentele sunt liganzi care afectează funcția proteinelor. Interacțiunea proteinelor cu liganzii este specifică. Cu toate acestea, datorită labilității conformaționale a proteinei și a centrului său activ, este posibil să se selecteze o altă substanță care ar putea interacționa și cu proteina din centrul activ sau din altă parte a moleculei.
Se numește o substanță similară ca structură cu un ligand natural analog structural al ligandului sau un ligand nenatural. De asemenea, interacționează cu proteina de la locul activ. Un analog structural al unui ligand poate îmbunătăți funcția proteinei (agonist),și reduceți-l (antagonist). Ligandul și analogii săi structurali concurează unul cu celălalt pentru legarea de proteină în același loc. Astfel de substanțe sunt numite modulatori competitivi(regulatori) ai funcţiilor proteinelor. Multe medicamente acționează ca inhibitori de proteine. Unele dintre ele sunt obținute prin modificarea chimică a liganzilor naturali. Inhibitorii funcțiilor proteinelor pot fi medicamente și otrăvuri.
Atropina este un inhibitor competitiv al receptorilor M-colinergici. Acetilcolina este un neurotransmitator pentru transmiterea impulsurilor nervoase prin sinapsele colinergice. Pentru a efectua excitația, acetilcolina eliberată în fanta sinaptică trebuie să interacționeze cu proteina receptoră a membranei postsinaptice. Două tipuri găsite receptori colinergici:
receptor M pe langa acetilcolina, interactioneaza selectiv cu muscarina (toxina agaric musca). M - receptorii colinergici sunt prezenți pe mușchii netezi și, atunci când interacționează cu acetilcolina, provoacă contracția acestora;
receptor H se leagă în mod specific de nicotină. Receptorii N-colinergici se găsesc la sinapsele mușchilor scheletici striați.
Inhibitor specific Receptorii M-colinergici este atropina. Se găsește în plantele belladonă și henbane.
 Atropina are grupe funcționale similare ca structură cu acetilcolina și aranjarea lor spațială, prin urmare este un inhibitor competitiv al receptorilor M-colinergici. Având în vedere că legarea acetilcolinei de receptorii M-colinergici determină contracția mușchilor netezi, atropina este utilizată ca medicament care ameliorează spasmul acestora. (antispasmodic). Astfel, este cunoscută utilizarea atropinei pentru a relaxa mușchii ochilor la vizualizarea fundului de ochi, precum și pentru a ameliora spasmele în timpul colicilor gastrointestinale. Receptorii M-colinergici sunt prezenți și în sistemul nervos central (SNC), astfel încât dozele mari de atropină pot provoca o reacție nedorită a sistemului nervos central: agitație motorie și psihică, halucinații, convulsii.
Atropina are grupe funcționale similare ca structură cu acetilcolina și aranjarea lor spațială, prin urmare este un inhibitor competitiv al receptorilor M-colinergici. Având în vedere că legarea acetilcolinei de receptorii M-colinergici determină contracția mușchilor netezi, atropina este utilizată ca medicament care ameliorează spasmul acestora. (antispasmodic). Astfel, este cunoscută utilizarea atropinei pentru a relaxa mușchii ochilor la vizualizarea fundului de ochi, precum și pentru a ameliora spasmele în timpul colicilor gastrointestinale. Receptorii M-colinergici sunt prezenți și în sistemul nervos central (SNC), astfel încât dozele mari de atropină pot provoca o reacție nedorită a sistemului nervos central: agitație motorie și psihică, halucinații, convulsii.
Ditilina este un agonist competitiv al receptorilor H-colinergici, inhibă funcția sinapselor neuromusculare.
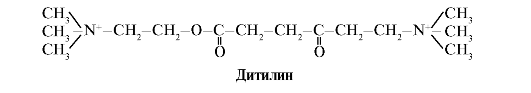 Sinapsele neuromusculare ale mușchilor scheletici conțin receptori H-colinergici. Interacțiunea lor cu acetilcolina duce la contracții musculare. În timpul unor operații chirurgicale, precum și în studiile endoscopice, sunt utilizate medicamente care provoacă relaxarea mușchilor scheletici. (relaxante musculare). Acestea includ ditilina, care este un analog structural al acetilcolinei. Se ataseaza de receptorii H-colinergici, dar spre deosebire de acetilcolina, este distrusa foarte lent de enzima acetilcolinesteraza. Ca urmare a deschiderii prelungite a canalelor ionice și a depolarizării persistente a membranei, conducerea impulsurilor nervoase este perturbată și are loc relaxarea musculară. Inițial, aceste proprietăți au fost descoperite în otravă de curare, motiv pentru care se numesc astfel de medicamente ca curare.
Sinapsele neuromusculare ale mușchilor scheletici conțin receptori H-colinergici. Interacțiunea lor cu acetilcolina duce la contracții musculare. În timpul unor operații chirurgicale, precum și în studiile endoscopice, sunt utilizate medicamente care provoacă relaxarea mușchilor scheletici. (relaxante musculare). Acestea includ ditilina, care este un analog structural al acetilcolinei. Se ataseaza de receptorii H-colinergici, dar spre deosebire de acetilcolina, este distrusa foarte lent de enzima acetilcolinesteraza. Ca urmare a deschiderii prelungite a canalelor ionice și a depolarizării persistente a membranei, conducerea impulsurilor nervoase este perturbată și are loc relaxarea musculară. Inițial, aceste proprietăți au fost descoperite în otravă de curare, motiv pentru care se numesc astfel de medicamente ca curare.
TEMA 1.3. DENATURAREA PROTEINELOR ŞI POSIBILITATEA RENATĂRII LOR SPONTANE
1. Deoarece conformația nativă a proteinelor este menținută din cauza interacțiunilor slabe, modificări ale compoziției și proprietăților mediului din jurul proteinei, expunerea la reactivi chimici și factori fizici provoacă o modificare a conformației acestora (proprietatea labilitatii conformaționale). Ruperea unui număr mare de legături duce la distrugerea conformației native și la denaturarea proteinelor.
Denaturarea proteinelor- aceasta este distrugerea conformatiei lor native sub influenta agentilor denaturanti, cauzata de ruperea legaturilor slabe care stabilizeaza structura spatiala a proteinei. Denaturarea este însoțită de distrugerea structurii unice tridimensionale și a centrului activ al proteinei și de pierderea activității sale biologice (Fig. 1.16).
Toate moleculele denaturate ale unei proteine dobândesc o conformație aleatorie care diferă de alte molecule ale aceleiași proteine. Radicalii de aminoacizi care formează centrul activ se dovedesc a fi distanțați spațial unul de celălalt, adică. locul de legare specific al proteinei cu ligand este distrus. În timpul denaturarii, structura primară a proteinelor rămâne neschimbată.
Aplicarea agenților de denaturare în cercetarea biologică și medicină.În studiile biochimice, înainte de a determina compușii cu greutate moleculară mică din materialul biologic, proteinele sunt de obicei îndepărtate mai întâi din soluție. Acidul tricloroacetic (TCA) este cel mai des folosit în acest scop. După adăugarea TCA la soluție, proteinele denaturate precipită și sunt ușor îndepărtate prin filtrare (Tabelul 1.1.)
În medicină, agenții de denaturare sunt adesea utilizați pentru sterilizarea instrumentelor și materialelor medicale în autoclave (agentul de denaturare este la temperatură ridicată) și ca antiseptice (alcool, fenol, cloramină) pentru tratarea suprafețelor contaminate care conțin microfloră patogenă.
2. Reactivarea proteinelor spontane- dovada determinismului structurii, conformaţiei şi funcţiei primare a proteinelor. Proteinele individuale sunt produse ale unei gene care au o secvență identică de aminoacizi și dobândesc aceeași conformație în celulă. Concluzia fundamentală că structura primară a unei proteine conține deja informații despre conformația și funcția sa a fost făcută pe baza capacității unor proteine (în special, ribonucleaza și mioglobina) de a se renativa spontan - restabili conformația lor nativă după denaturare.
Formarea structurilor proteice spațiale se realizează prin metoda auto-asamblarii - un proces spontan în care un lanț polipeptidic, care are o structură primară unică, tinde să adopte o conformație cu cea mai scăzută energie liberă în soluție. Capacitatea de a renativa proteinele care își păstrează structura primară după denaturare a fost descrisă într-un experiment cu enzima ribonuclează.
Ribonucleaza este o enzimă care descompune legăturile dintre nucleotidele individuale dintr-o moleculă de ARN. Această proteină globulară are un lanț polipeptidic, a cărui structură terțiară este stabilizată de multe legături slabe și patru legături disulfurice.
Tratamentul ribonucleazei cu uree, care rupe legăturile de hidrogen din moleculă și un agent reducător, care rupe legăturile disulfurice, duce la denaturarea enzimei și la pierderea activității acesteia.
Îndepărtarea agenților denaturanți prin dializă duce la restabilirea conformației și funcției proteinelor, de exemplu. a renaște. (Fig. 1.17).
 Orez. 1.17. Denaturarea și renativarea ribonucleazei
Orez. 1.17. Denaturarea și renativarea ribonucleazei
A - conformația nativă a ribonucleazei, în structura terțiară a cărei patru legături disulfurice; B - moleculă de ribonuclează denaturată;
B - moleculă de ribonuclează reactivată cu structură și funcție restaurate
1. Completați tabelul 1.2.
Tabelul 1.2. Clasificarea aminoacizilor în funcție de polaritatea radicalilor
2. Scrieți formula tetrapeptidei:
Asp - Pro - Fen - Liz
a) evidențiați grupările repetate din peptidă care formează coloana vertebrală peptidică și grupările variabile reprezentate de radicalii aminoacizi;
b) etichetați N- și C-terminali;
c) evidenţiază legăturile peptidice;
d) scrieți o altă peptidă formată din aceiași aminoacizi;
e) numărați numărul de variante posibile ale unei tetrapeptide cu o compoziție similară de aminoacizi.
3. Explicați rolul structurii primare a proteinelor folosind exemplul unei analize comparative a doi hormoni peptidici similari din punct de vedere structural și apropiați din punct de vedere evolutiv ai neurohipofizei mamiferelor - oxitocina și vasopresina (Tabelul 1.3).
Tabelul 1.3. Structura și funcțiile oxitocinei și vasopresinei
 Pentru aceasta:
Pentru aceasta:
a) comparați compoziția și secvența de aminoacizi a două peptide;
b) găsiți asemănarea structurii primare a celor două peptide și asemănarea acțiunii lor biologice;
c) găsiți diferențe în structura a două peptide și diferențe în funcțiile acestora;
d) trageți o concluzie despre influența structurii primare a peptidelor asupra funcțiilor acestora.
4. Descrieți principalele etape de formare a conformației proteinelor globulare (structuri secundare, terțiare, conceptul de structură supersecundară). Indicați tipurile de legături implicate în formarea structurilor proteice. Ce radicali de aminoacizi pot participa la formarea interacțiunilor hidrofobe, ionice, legături de hidrogen.
Dă exemple.
5. Definiți conceptul de „labilitatea conformațională a proteinelor”, indicați motivele existenței și semnificația acestuia.
6. Extindeți semnificația următoarei fraze: „Funcționarea proteinelor se bazează pe interacțiunea lor specifică cu ligandul”, folosind termenii și explicând sensul lor: conformație proteinei, centru activ, ligand, complementaritate, funcție proteică.
7. Folosind un exemplu, explicați ce domenii sunt și care este rolul lor în funcționarea proteinelor.
SARCINI DE AUTOCONTROL
1. Meci.
Grupa funcțională în radicalul aminoacid:
A. Gruparea carboxil B. Gruparea hidroxil C Gruparea guanidină D. Gruparea tiol E. Gruparea amino
2. Alege raspunsurile corecte.
Aminoacizii cu radicali polari neîncărcați sunt:
A. Cis B. Asn
B. Glu G. Trei
3. Alege raspunsurile corecte.
Radicali de aminoacizi:
A. Asigură specificitatea structurii primare B. Participa la formarea structurii terțiare
B. Situate pe suprafața proteinei, ele influențează solubilitatea acesteia D. Formează centrul activ
D. Participa la formarea legăturilor peptidice
4. Alege raspunsurile corecte.
Interacțiunile hidrofobe se pot forma între radicalii de aminoacizi:
A. Tre Lay B. Pro Trei
B. Met Ile G. Tir Ala D. Val Fen
5. Alege raspunsurile corecte.
Între radicalii de aminoacizi se pot forma legături ionice:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn apr
6. Alege raspunsurile corecte.
Legăturile de hidrogen se pot forma între radicalii de aminoacizi:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Meci.
Tipul de legătură implicat în formarea structurii proteinei:
A. Structura primară B. Structura secundară
B. Structura terţiară
D. Structura suprasecundară E. Conformaţia.
1. Legături de hidrogen între atomii scheletului peptidic
2. Legături slabe între grupările funcționale ale radicalilor aminoacizi
3. Legături între grupările α-amino și α-carboxil ale aminoacizilor
8. Alege raspunsurile corecte. Tripsina:
A. Enzima proteolitică B. Conține două domenii
B. Hidrolizează amidonul
D. Site-ul activ este situat între domenii. D. Constă din două lanțuri polipeptidice.
9. Alege raspunsurile corecte. Atropina:
A. Neurotransmițător
B. Analog structural al acetilcolinei
B. Interacționează cu receptorii H-colinergici
D. Intareste conducerea impulsurilor nervoase prin sinapsele colinergice
D. Inhibitor competitiv al receptorilor M-colinergici
10. Alegeți afirmațiile corecte. În proteine:
A. Structura primară conține informații despre structura site-ului său activ
B. Centrul activ se formează la nivelul structurii primare
B. Conformația este fixată rigid prin legături covalente
D. Situl activ poate interacționa cu un grup de liganzi similari
datorită labilităţii conformaţionale a proteinelor D. Modificările mediului pot afecta afinitatea activului
centru la ligand
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
TERMENI ȘI CONCEPTE DE BAZĂ
1. Proteine, polipeptide, aminoacizi
2. Structuri proteice primare, secundare, terțiare
3. Conformația, conformația proteinei native
4. Legături covalente și slabe în proteine
5. Labilitatea conformațională
6. Sit activ proteic
7. Liganzi
8. Plierea proteinelor
9. Analogi structurali ai liganzilor
10. Proteine de domeniu
11. Proteine simple și complexe
12. Denaturarea proteinelor, agenți de denaturare
13. Reactivarea proteinelor
Rezolva probleme
„Organizarea structurală a proteinelor și baza funcționării lor”
1. Funcția principală a proteinei - hemoglobina A (HbA) este transportul oxigenului către țesuturi. În populația umană, sunt cunoscute forme multiple ale acestei proteine cu proprietăți și funcții modificate - așa-numitele hemoglobine anormale. De exemplu, hemoglobina S, găsită în globulele roșii ale pacienților cu siclemie (HbS), s-a dovedit a avea o solubilitate scăzută în condiții de presiune parțială scăzută a oxigenului (cum este cazul sângelui venos). Acest lucru duce la formarea de agregate ale acestei proteine. Proteina își pierde funcția, precipită, iar celulele roșii din sânge devin neregulate (unele dintre ele formează o formă de seceră) și sunt distruse mai repede decât de obicei în splină. Ca urmare, se dezvoltă anemia cu celule falciforme.
Singura diferență în structura primară a HbA a fost găsită în regiunea N-terminală a lanțului β al hemoglobinei. Comparați regiunile N-terminale ale catenei β și arată cum modificările în structura primară a proteinei îi afectează proprietățile și funcțiile.
 Pentru aceasta:
Pentru aceasta:
a) scrieți formulele aminoacizilor prin care HbA diferă și comparați proprietățile acestor aminoacizi (polaritate, sarcină).
b) trageți o concluzie despre motivul scăderii solubilității și perturbării transportului oxigenului în țesuturi.
2. Figura prezintă o diagramă a structurii unei proteine care are un centru de legare cu un ligand (centru activ). Explicați de ce proteina este selectivă în alegerea ligandului. Pentru aceasta:
a) amintiți-vă care este centrul activ al unei proteine și luați în considerare structura centrului activ al proteinei prezentată în figură;
b) scrieți formulele radicalilor aminoacizi care alcătuiesc centrul activ;
c) trageți un ligand care ar putea interacționa în mod specific cu situsul activ al proteinei. Indicați pe el grupele funcționale care pot forma legături cu radicalii aminoacizi care alcătuiesc centrul activ;
d) indicați tipurile de legături care apar între radicalii ligand și aminoacizi ai situsului activ;
e) explicați pe ce se bazează specificitatea interacțiunii proteină-ligand.
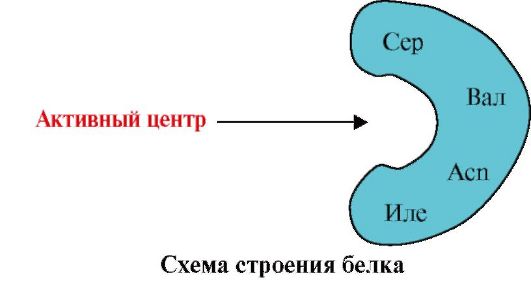 3.
Figura prezintă locul activ al proteinei și al mai multor liganzi.
3.
Figura prezintă locul activ al proteinei și al mai multor liganzi.
Determinați care ligand este cel mai probabil să interacționeze cu situsul activ al proteinei și de ce.
 Ce tipuri de legături apar în timpul formării unui complex proteină-ligand?
Ce tipuri de legături apar în timpul formării unui complex proteină-ligand?
4. Analogii structurali ai liganzilor proteici naturali pot fi utilizați ca medicamente pentru a modifica activitatea proteinelor.
Acetilcolina este un mediator al transmiterii excitației la sinapsele neuromusculare. Când acetilcolina interacționează cu proteinele - receptorii membranei postsinaptice a mușchilor scheletici, canalele ionice se deschid și are loc contracția musculară. Ditilin este un medicament folosit în unele operații pentru relaxarea mușchilor, deoarece întrerupe transmiterea impulsurilor nervoase prin sinapsele neuromusculare. Explicați mecanismul de acțiune al ditilinei ca relaxant muscular. Pentru aceasta:
a) scrieți formulele acetilcolinei și ditilinei și comparați structurile acestora;
b) descrieți mecanismul efectului relaxant al ditilinei.
5. În unele boli, temperatura corpului pacientului crește, ceea ce este considerat o reacție de protecție a corpului. Cu toate acestea, temperaturile ridicate sunt dăunătoare pentru proteinele corpului. Explicați de ce la temperaturi peste 40 °C funcția proteinelor este perturbată și apare o amenințare la adresa vieții umane. Pentru a face acest lucru, amintiți-vă:
1) Structura proteinelor și legăturile care îi mențin structura în conformația nativă;
2) Cum se modifică structura și funcția proteinelor odată cu creșterea temperaturii?;
3) Ce este homeostazia și de ce este importantă pentru menținerea sănătății umane.
Unitate modulară 2 PROTEINE OLIGOMERICE CA ȚINTE ALE INFLUENȚELOR REGLATORII. DIVERSITATEA STRUCTURALA SI FUNCTIONALA A PROTEINELOR. METODE DE SEPARARE ȘI PURIFICARE A PROTEINELOR
Obiective de învățare A fi capabil să:
1. Utilizați cunoștințele despre caracteristicile structurii și funcțiilor proteinelor oligomerice pentru a înțelege mecanismele adaptative de reglare a funcțiilor acestora.
2. Explicați rolul chaperonelor în sinteza și menținerea conformației proteinelor în condiții celulare.
3. Explicați varietatea manifestărilor vieții prin varietatea structurilor și funcțiilor proteinelor sintetizate în organism.
4. Analizați relația dintre structura proteinelor și funcția lor folosind exemple de comparare a hemoproteinelor înrudite - mioglobină și hemoglobină, precum și reprezentanți ai celor cinci clase de proteine din familia imunoglobulinelor.
5. Aplicarea cunoștințelor despre particularitățile proprietăților fizice și chimice ale proteinelor pentru a selecta metode de purificare a acestora din alte proteine și impurități.
6. Interpretați rezultatele compoziției cantitative și calitative a proteinelor plasmatice pentru a confirma sau clarifica diagnosticul clinic.
Știi:
1. Caracteristici ale structurii proteinelor oligomerice și mecanisme adaptative de reglare a funcțiilor acestora folosind exemplul hemoglobinei.
2. Structura și funcțiile chaperonelor și importanța acestora pentru menținerea conformației native a proteinelor în condiții celulare.
3. Principii de combinare a proteinelor în familii pe baza asemănării conformației și funcțiilor lor folosind exemplul imunoglobulinelor.
4. Metode de separare a proteinelor pe baza caracteristicilor proprietăților lor fizico-chimice.
5. Electroforeza plasmei sanguine ca metodă de evaluare a compoziției calitative și cantitative a proteinelor.
TEMA 1.4. CARACTERISTICI ALE STRUCTURII ȘI FUNCȚIONĂRII PROTEINELOR OLIGOMERICE CU EXEMPLU DE HEMOGLOBINĂ
1. Multe proteine conțin mai multe lanțuri polipeptidice. Astfel de proteine sunt numite oligomerice,și lanțuri individuale - protomeri. Protomerii din proteinele oligomerice sunt legați prin multe legături slabe necovalente (hidrofobe, ionice, hidrogen). Interacţiune
protomerii se realizează datorită complementaritatea suprafețele lor de contact.
Numărul de protomeri din proteinele oligomerice poate varia foarte mult: hemoglobina conține 4 protomeri, enzima aspartat aminotransferaza are 12 protomeri, iar proteina virusului mozaicului tutunului conține 2120 protomeri legați prin legături necovalente. În consecință, proteinele oligomerice pot avea greutăți moleculare foarte mari.
Interacțiunea unui protomer cu alții poate fi considerată un caz special de interacțiune proteină-ligand, deoarece fiecare protomer servește ca ligand pentru alți protomeri. Numărul și metoda de unire a protomerilor într-o proteină se numesc structura proteinelor cuaternare.
Proteinele pot conține protomeri cu structuri identice sau diferite, de exemplu, homodimerii sunt proteine care conțin doi protomeri identici, iar heterodimerii sunt proteine care conțin doi protomeri diferiți.
Dacă proteinele conțin protomeri diferiți, atunci se pot forma pe ele centre de legare cu liganzi diferiți, care diferă ca structură. Când un ligand se leagă de situsul activ, se manifestă funcția acestei proteine. Un centru situat pe un protomer diferit se numește alosteric (diferit de cel activ). Contactarea ligand sau efector alosteric,îndeplineşte o funcţie de reglementare (Fig. 1.18). Interacțiunea centrului alosteric cu efectorul provoacă modificări conformaționale în structura întregii proteine oligomerice datorită labilității conformaționale a acesteia. Acest lucru afectează afinitatea situsului activ pentru un ligand specific și reglează funcția acelei proteine. O schimbare în conformația și funcția tuturor protomerilor în timpul interacțiunii unei proteine oligomere cu cel puțin un ligand se numește modificări conformaționale cooperative. Se numesc efectori care îmbunătățesc funcția proteinelor activatori,și efectori care îi inhibă funcția - inhibitori.
Astfel, proteinele oligomerice, precum și proteinele cu structură de domeniu, au o nouă proprietate în comparație cu proteinele monomerice - capacitatea de a regla alosteric funcțiile (reglare prin atașarea diferiților liganzi la proteină). Acest lucru poate fi văzut comparând structurile și funcțiile a două proteine complexe strâns legate, mioglobina și hemoglobina.
 Orez. 1.18. Schema structurii unei proteine dimerice
Orez. 1.18. Schema structurii unei proteine dimerice
2. Formarea structurilor spațiale și funcționarea mioglobinei.
Mioglobina (Mb) este o proteină care se găsește în mușchii roșii, a cărei funcție principală este de a crea rezerve de O 2 necesare lucrului muscular intens. Mb este o proteină complexă care conține o parte proteică - apoMb și o parte neproteică - hem. Structura primară a apoMB determină conformația sa globulară compactă și structura centrului activ, de care este atașată partea neproteică a mioglobinei, hem. Oxigenul care vine din sânge către mușchi se leagă de hemii Fe+2 din mioglobină. Mb este o proteină monomerică care are o afinitate foarte mare pentru O 2, prin urmare eliberarea de oxigen de către mioglobină are loc numai în timpul muncii musculare intense, când presiunea parțială a O 2 scade brusc.
Formarea conformaţiei Mv.În mușchii roșii, pe ribozomi în timpul translației, se sintetizează structura primară a MB, reprezentată de o secvență specifică de 153 de resturi de aminoacizi. Structura secundară a lui Mb conține opt elice α, numite cu litere latine de la A la H, între care există regiuni nehelicoidale. Structura terțiară a lui Mb are forma unui globul compact, în adâncitura căruia se află centrul activ între elicele α F și E (Fig. 1.19).
 Orez. 1.19. Structura mioglobinei
Orez. 1.19. Structura mioglobinei
3. Caracteristici ale structurii și funcționării centrului activ al MV. Centrul activ al Mb este format predominant din radicali de aminoacizi hidrofobi, distanțați larg unul de celălalt în structura primară (de exemplu, Tri 3 9 și Fen 138) Puțin solubili în apă liganzii - hem și O 2 - se atașează la centrul activ. Hemul este un ligand specific al apoMB (Fig. 1.20), a cărui bază este alcătuită din patru inele pirol conectate prin punți metanilice; în centru se află un atom de Fe+ 2 legat de atomii de azot ai inelelor pirol prin patru legături de coordonare. În centrul activ al Mb, pe lângă radicalii de aminoacizi hidrofobi, există și reziduuri a doi aminoacizi cu radicali hidrofili - Gis E 7(Gis 64) și GIS F 8(His 93) (Fig. 1.21).
 Orez. 1.20. Structura hemului - partea neproteică a mioglobinei și hemoglobinei
Orez. 1.20. Structura hemului - partea neproteică a mioglobinei și hemoglobinei
 Orez. 1.21. Localizarea hemului și a O2 în locul activ al apomioglobinei și al protomerilor hemoglobinei
Orez. 1.21. Localizarea hemului și a O2 în locul activ al apomioglobinei și al protomerilor hemoglobinei
Hemul este legat covalent de His F8 printr-un atom de fier. O 2 se atașează de fierul de călcat pe cealaltă parte a planului hem. Lui E 7 este necesar pentru orientarea corectă a O 2 și facilitează adăugarea de oxigen la Fe + 2 hem
GIS F 8 formează o legătură de coordonare cu Fe+ 2 și fixează ferm hemul în centrul activ. Gis E 7 necesar pentru orientarea corectă în centrul activ al altui ligand - O 2 în timpul interacțiunii acestuia cu Fe + 2 hem. Micromediul hemului creează condiții pentru legarea puternică, dar reversibilă, a O2 de Fe + 2 și împiedică intrarea apei în locul activ hidrofob, ceea ce poate duce la oxidarea acestuia la Fe + 3.
Structura monomerică a Mb și centrul său activ determină afinitatea ridicată a proteinei pentru O2.
4. Structura oligomerică a Hb și reglarea afinității Hb față de liganzii O2. Hemoglobinele umane- o familie de proteine, precum mioglobina, legate de proteine complexe (hemoproteine). Au o structură tetramerică și conțin două lanțuri α, dar diferă prin structura celorlalte două lanțuri polipeptidice (lanțuri 2α, 2x). Structura celui de-al doilea lanț polipeptidic determină caracteristicile funcționării acestor forme de Hb. Aproximativ 98% din hemoglobina din globulele roșii ale unui adult este hemoglobina A(2α-, 2p-lanțuri).
În timpul dezvoltării fetale, funcționează două tipuri principale de hemoglobine: Hb embrionară(2α, 2ε), care se găsește în stadiile incipiente ale dezvoltării fetale și hemoglobina F (fetală)- (2α, 2γ), care înlocuiește hemoglobina fetală precoce în luna a șasea de dezvoltare intrauterină și numai după naștere este înlocuită cu Hb A.
HB A este o proteină legată de mioglobina (MB) găsită în globulele roșii umane adulte. Structura protomerilor săi individuali este similară cu cea a mioglobinei. Structurile secundare și terțiare ale mioglobinei și protomerilor hemoglobinei sunt foarte asemănătoare, în ciuda faptului că în structura primară a lanțurilor lor polipeptidice doar 24 de resturi de aminoacizi sunt identice (structura secundară a protomerilor hemoglobinei, ca și mioglobina, conține opt α-helice, desemnată prin litere latine de la A la H , iar structura terţiară are forma unui globule compact). Dar, spre deosebire de mioglobina, hemoglobina are o structură oligomerică, constând din patru lanțuri polipeptidice conectate prin legături necovalente (Figura 1.22).
Fiecare protomer Hb este asociat cu o parte non-proteică - hem și protomeri vecini. Legătura părții proteice a Hb cu hem este similară cu cea a mioglobinei: în centrul activ al proteinei, părțile hidrofobe ale hemului sunt înconjurate de radicali hidrofobi de aminoacizi, cu excepția His F 8 și His E 7, care sunt situate pe ambele părți ale planului hem și joacă un rol similar în funcționarea proteinei și în legarea acesteia cu oxigenul (vezi structura mioglobinei).
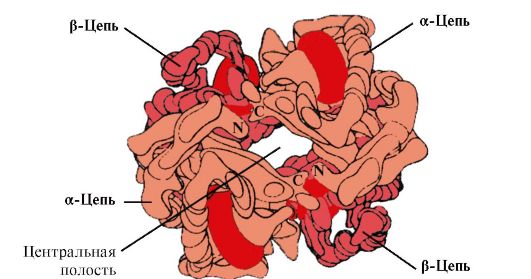 Orez. 1.22. Structura oligomerică a hemoglobinei
Orez. 1.22. Structura oligomerică a hemoglobinei
In afara de asta, Gis E 7 realizează un important rol suplimentarîn funcționarea Nv. Hemul liber are o afinitate de 25.000 de ori mai mare pentru CO decât pentru O2. CO se formează în cantități mici în organism și, având în vedere afinitatea sa mare pentru hem, ar putea perturba transportul de O 2 necesar vieții celulare. Totuși, în compoziția hemoglobinei, afinitatea hemului pentru monoxidul de carbon depășește de numai 200 de ori afinitatea pentru O 2 datorită prezenței His E 7 în centrul activ. Restul acestui aminoacid creează condiții optime pentru legarea hemului de O2 și slăbește interacțiunea hemului cu CO.
5. Funcția principală a HB este transportul O2 de la plămâni la țesuturi. Spre deosebire de mioglobina monomerică, care are o afinitate foarte mare pentru O2 și îndeplinește funcția de stocare a oxigenului în mușchii roșii, structura oligomerică a hemoglobinei oferă:
1) saturarea rapidă a HB cu oxigen în plămâni;
2) capacitatea HB de a elibera oxigen în ţesuturi la o presiune parţială relativ mare de O2 (20-40 mm Hg);
3) posibilitatea de reglare a afinității Hb față de O 2.
6. Modificările cooperante în conformația protomerilor hemoglobinei accelerează legarea O 2 în plămâni și eliberarea acestuia în țesuturi. În plămâni, presiunea parțială ridicată a O2 promovează legarea acestuia de Hb în situsul activ a patru protomeri (2α și 2β). Centrul activ al fiecărui protomer, ca și în mioglobină, este situat între două elice α (F și E) într-un buzunar hidrofob. Conține o parte non-proteică - hem, atașată de partea proteică prin multe interacțiuni hidrofobe slabe și o legătură puternică între Fe 2 + hem și His F 8 (vezi Fig. 1.21).
În deoxihemoglobină, datorită acestei legături cu His F 8, atomul de Fe 2 + iese din planul hem către histidină. Legarea O2 la Fe2+ are loc pe cealaltă parte a hemului în regiunea His E7 folosind o singură legătură de coordonare liberă. His E 7 oferă condiții optime pentru legarea O 2 de fierul hem.
Adăugarea de O 2 la atomul de Fe + 2 al unui protomer determină mișcarea acestuia în planul hem, urmată de reziduul de histidină asociat cu acesta
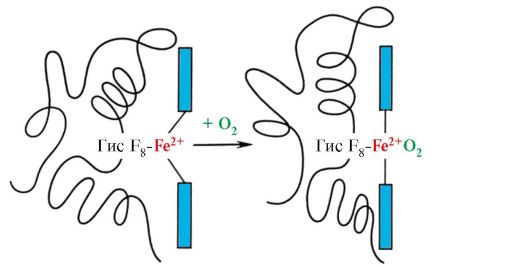 Orez. 1.23. Modificarea conformației protomerului hemoglobinei atunci când este combinat cu O2
Orez. 1.23. Modificarea conformației protomerului hemoglobinei atunci când este combinat cu O2
Acest lucru duce la o modificare a conformației tuturor lanțurilor polipeptidice datorită labilității lor conformaționale. Modificarea conformației altor lanțuri facilitează interacțiunea acestora cu moleculele de O 2 ulterioare.
A patra moleculă de O 2 se atașează de hemoglobină de 300 de ori mai ușor decât prima (Fig. 1.24).
 Orez. 1.24. Modificări cooperante în conformația protomerilor hemoglobinei în timpul interacțiunii sale cu O2
Orez. 1.24. Modificări cooperante în conformația protomerilor hemoglobinei în timpul interacțiunii sale cu O2
În țesuturi, fiecare moleculă de O2 ulterioară este scindată mai ușor decât cea anterioară, tot datorită modificărilor cooperante în conformația protomerilor.
7. CO 2 şi H+, formate în timpul catabolismului substanţelor organice, reduc afinitatea hemoglobinei pentru O 2 proporţional cu concentraţia acestora. Energia necesară pentru funcționarea celulelor este produsă în primul rând în mitocondrii în timpul oxidării substanțelor organice folosind O 2 eliberat din plămâni de hemoglobină. Ca urmare a oxidării substanţelor organice se formează produşii finali ai descompunerii acestora: CO 2 şi K 2 O, a căror cantitate este proporţională cu intensitatea proceselor de oxidare în curs.
CO 2 difuzează din celule în sânge și pătrunde în celulele roșii din sânge, unde, sub acțiunea enzimei carbanhidraze, este transformat în acid carbonic. Acest acid slab se disociază într-un proton și un ion de bicarbonat.
H+ sunt capabili să se alăture radicalilor Săi 14 6 în lanțurile α și β ale hemoglobinei, adică în zone îndepărtate de hem. Protonarea hemoglobinei reduce afinitatea acesteia pentru O 2, favorizează eliminarea O 2 din oxiHb, formarea deoxiHb și crește aportul de oxigen către țesuturi proporțional cu numărul de protoni formați (Fig. 1.25).
O creștere a cantității de oxigen eliberat în funcție de creșterea concentrației de H+ în celulele roșii din sânge se numește efect Bohr (numit după fiziologul danez Christian Bohr, care a descoperit primul acest efect).
În plămâni, o presiune parțială mare a oxigenului promovează legarea acestuia la deoxiHb, ceea ce reduce afinitatea proteinei pentru H +. Protonii eliberați sub acțiunea acidului carbonic reacționează cu bicarbonații pentru a forma CO 2 și H 2 O

 Orez. 1.25. Dependența afinității Hb pentru O 2 de concentrația de CO 2 și protoni (efectul Bohr):
Orez. 1.25. Dependența afinității Hb pentru O 2 de concentrația de CO 2 și protoni (efectul Bohr):
A- influenţa concentraţiei de CO 2 şi H+ asupra eliberării de O 2 din complexul cu HB (efect Bohr); B- oxigenarea deoxihemoglobinei în plămâni, formarea și eliberarea de CO2.
CO2 rezultat intră în spațiul alveolar și este îndepărtat cu aerul expirat. Astfel, cantitatea de oxigen eliberată de hemoglobină în țesuturi este reglată de produșii catabolismului substanțelor organice: cu cât descompunerea substanțelor este mai intensă, de exemplu în timpul exercițiului fizic, cu atât concentrația de CO 2 și H + este mai mare și cu atât mai mult oxigen. ţesuturile primesc ca urmare a scăderii afinităţii Hb pentru O 2.
8. Reglarea alosterică a afinității Hb pentru O2 de către ligand - 2,3-bisfosfoglicerat.În eritrocite, ligandul alosteric al hemoglobinei, 2,3-bisfosfogliceratul (2,3-BPG), este sintetizat din produsul oxidării glucozei - 1,3-bisfosfoglicerat. În condiții normale, concentrația de 2,3-BPG este ridicată și comparabilă cu concentrația de Hb. 2,3-BPG are o sarcină negativă puternică de -5.
 Bisfosfogliceratul din capilarele tisulare, care se leagă de deoxihemoglobină, crește eliberarea de oxigen în țesuturi, reducând afinitatea Hb pentru O2.
Bisfosfogliceratul din capilarele tisulare, care se leagă de deoxihemoglobină, crește eliberarea de oxigen în țesuturi, reducând afinitatea Hb pentru O2.
În centrul moleculei de hemoglobină tetramerică se află o cavitate. Este format din resturile de aminoacizi ale tuturor celor patru protomeri (vezi Fig. 1.22). În capilarele tisulare, protonarea Hb (efectul Bohr) duce la ruperea legăturii dintre fierul hem și O2. Într-o moleculă
deoxihemoglobina, comparativ cu oxihemoglobina, apar legături ionice suplimentare care leagă protomerii, drept urmare dimensiunile cavității centrale cresc față de oxihemoglobina. Cavitatea centrală este locul de atașare a 2,3-BPG la hemoglobină. Datorită diferenței de dimensiune a cavității centrale, 2,3-BPG se poate atașa doar la deoxihemoglobină.
2,3-BPG interacționează cu hemoglobina într-un loc îndepărtat de centrii activi ai proteinei și aparține alosterică liganzi (reglatori), iar cavitatea centrală a Hb este centru alosteric. 2,3-BPG are o sarcină negativă puternică și interacționează cu cinci grupe încărcate pozitiv ale celor două lanțuri β ale Hb: gruparea α-amino N-terminală a lui Val și radicalii Lys 82 His 143 (Fig. 1.26).
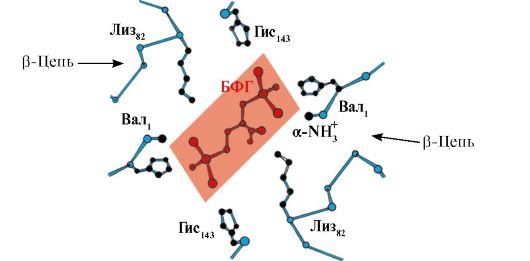 Orez. 1.26. BPG în cavitatea centrală a deoxihemoglobinei
Orez. 1.26. BPG în cavitatea centrală a deoxihemoglobinei
BPG se leagă de trei grupuri încărcate pozitiv pe fiecare catenă β.
În capilarele tisulare, deoxihemoglobina rezultată interacționează cu 2,3-BPG și se formează legături ionice între radicalii încărcați pozitiv ai lanțurilor β și ligandul încărcat negativ, care modifică conformația proteinei și reduc afinitatea Hb pentru O2. . O scădere a afinității Hb pentru O 2 contribuie la o eliberare mai eficientă a O 2 în țesut.
În plămâni, la presiune parțială mare, oxigenul interacționează cu Hb, unind fierul hem; în acest caz, conformația proteinei se modifică, cavitatea centrală scade și 2,3-BPG este deplasat din centrul alosteric.
Astfel, proteinele oligomerice au proprietăți noi în comparație cu proteinele monomerice. Atașarea liganzilor la situsuri
distanță spațial unul de celălalt (alosteric), poate provoca modificări conformaționale în întreaga moleculă proteică. Datorită interacțiunii cu liganzii de reglare, are loc o modificare a conformației și adaptarea funcției moleculei proteice la schimbările de mediu.
TEMA 1.5. MENȚINEREA CONFORMĂȚII NATIVE A PROTEINELOR ÎN CONDIȚII CELULARE
În celule, în timpul sintezei lanțurilor polipeptidice, transportul acestora prin membrane către părțile corespunzătoare ale celulei, în timpul procesului de pliere (formarea conformației native) și în timpul asamblării proteinelor oligomerice, precum și în timpul funcționării lor, intermediare. În structura proteinei apar conformații instabile, predispuse la agregare. Radicalii hidrofobi, de obicei ascunși în interiorul moleculei proteice în conformația nativă, apar la suprafață într-o conformație instabilă și tind să se combine cu grupuri de alte proteine care sunt slab solubile în apă. În celulele tuturor organismelor cunoscute, s-au găsit proteine speciale care asigură plierea optimă a proteinelor celulare, stabilizează conformația lor nativă în timpul funcționării și, cel mai important, mențin structura și funcțiile proteinelor intracelulare atunci când homeostazia este perturbată. Aceste proteine sunt numite "socitori" care înseamnă „dădacă” în franceză.
1. Chaperonele moleculare și rolul lor în prevenirea denaturarii proteinelor.
Chaperonele (CH) sunt clasificate în funcție de masa subunităților lor. Chaperonele cu greutate moleculară mare au o masă de la 60 la 110 kDa. Dintre acestea, trei clase au fost cele mai studiate: Sh-60, Sh-70 și Sh-90. Fiecare clasă include o familie de proteine înrudite. Astfel, Sh-70 include proteine cu o greutate moleculară de la 66 la 78 kDa. Chaperonele cu greutate moleculară mică au o greutate moleculară de la 40 la 15 kDa.
Printre însoțitori se numără constitutiv proteine, a căror sinteză bazală ridicată nu depinde de efectele stresului asupra celulelor organismului și inductibil, a cărui sinteză în condiții normale este slabă, dar crește brusc sub stres. Chaperonele inductibile sunt numite și „proteine de șoc termic”, deoarece au fost descoperite pentru prima dată în celulele expuse la temperaturi ridicate. În celule, datorită concentrației mari de proteine, reactivarea spontană a proteinelor parțial denaturate este dificilă. Sh-70 poate preveni apariția denaturarii și poate ajuta la restabilirea conformației native a proteinelor. Însoțitori moleculari-70- o clasă foarte conservată de proteine găsite în toate părțile celulei: citoplasmă, nucleu, reticul endoplasmatic, mitocondrii. La capătul carboxil al lanțului polipeptidic unic Ш-70 există o regiune care este un șanț capabil să interacționeze cu peptide de lungime
de la 7 la 9 resturi de aminoacizi îmbogățite cu radicali hidrofobi. Astfel de regiuni din proteinele globulare apar aproximativ la fiecare 16 aminoacizi. Sh-70 este capabil să protejeze proteinele de inactivarea temperaturii și să restabilească conformația și activitatea proteinelor parțial denaturate.
2. Rolul chaperonelor în plierea proteinelor.În timpul sintezei proteinelor pe ribozom, regiunea N-terminală a polipeptidei este sintetizată înaintea celei C-terminale. Pentru a forma conformația nativă, este necesară secvența completă de aminoacizi a proteinei. În procesul de sinteză a proteinelor, chaperonele-70, datorită structurii centrului lor activ, sunt capabile să închidă zonele polipeptidei predispuse la agregare, îmbogățite în radicali de aminoacizi hidrofobi până la finalizarea sintezei (Figura 1.27, A). ).
 Orez. 1.27. Participarea chaperonelor la plierea proteinelor
Orez. 1.27. Participarea chaperonelor la plierea proteinelor
A - participarea chaperonelor-70 la prevenirea interacțiunilor hidrofobe între secțiuni ale polipeptidei sintetizate; B - formarea conformației native a proteinei din complexul de chaperonă
Multe proteine cu molecule înalte care au o conformație complexă, cum ar fi o structură de domeniu, se pliază într-un spațiu special format din Sh-60. Ш-60 funcţionează ca un complex oligomeric format din 14 subunităţi. Ele formează două inele goale, fiecare dintre ele constând din șapte subunități, aceste inele sunt conectate între ele. Fiecare subunitate Sh-60 este formată din trei domenii: apical (apical), îmbogățit cu radicali hidrofobi orientați spre cavitatea inelului, intermediar și ecuatorial (Fig. 1.28).
 Orez. 1.28. Structura complexului de chaperonine constând din 14 Ш-60
Orez. 1.28. Structura complexului de chaperonine constând din 14 Ш-60
A - vedere laterală; B - vedere de sus
Proteinele sintetizate, care au elemente pe suprafața caracteristice moleculelor desfășurate, în special radicalii hidrofobi, intră în cavitatea inelelor chaperone. În mediul specific acestor cavități se caută conformații posibile până când se găsește singura care este cea mai favorabilă energetic (Fig. 1.27, B). Formarea conformațiilor și eliberarea proteinelor este însoțită de hidroliza ATP în regiunea ecuatorială. De obicei, o astfel de pliere dependentă de însoțitor necesită o cantitate semnificativă de energie.
Pe lângă participarea la formarea structurii tridimensionale a proteinelor și la renativarea proteinelor parțial denaturate, însoțitorii sunt, de asemenea, necesari pentru apariția unor procese fundamentale precum asamblarea proteinelor oligomerice, recunoașterea și transportul proteinelor denaturate în lizozomi, transportul proteinelor prin membrane și participarea la reglarea activității complexelor proteice.
TEMA 1.6. VARIETATE DE PROTEINE. FAMILIILE DE PROTEINE: EXEMPLU DE IMUNOGLOBULINE
1. Proteinele joacă un rol decisiv în viața celulelor individuale și a întregului organism multicelular, iar funcțiile lor sunt surprinzător de diverse. Acest lucru este determinat de caracteristicile structurii primare și conformațiilor proteinelor, de structura unică a centrului activ și de capacitatea de a lega liganzi specifici.
Doar o fracțiune foarte mică din toate variantele posibile de lanțuri peptidice poate adopta o structură spațială stabilă; majoritate
dintre ele pot lua multe conformații cu aproximativ aceeași energie Gibbs, dar cu proprietăți diferite. Structura primară a majorității proteinelor cunoscute selectate prin evoluția biologică asigură o stabilitate excepțională a uneia dintre conformații, ceea ce determină caracteristicile funcționării acestei proteine.
2. Familii de proteine.În cadrul aceleiași specii biologice, substituțiile de resturi de aminoacizi pot duce la apariția diferitelor proteine care îndeplinesc funcții înrudite și au secvențe de aminoacizi omoloage. Astfel de proteine înrudite au conformații surprinzător de similare: numărul și pozițiile relative ale elicelor a și/sau structurilor p și majoritatea spirelor și curburilor lanțurilor polipeptidice sunt similare sau identice. Proteinele cu regiuni omoloage ale lanțului polipeptidic, conformație similară și funcții înrudite sunt clasificate în familii de proteine. Exemple de familii de proteine: serin proteinaze, familia imunoglobulinelor, familia mioglobinelor.
Serin proteinaze- o familie de proteine care îndeplinesc funcția de enzime proteolitice. Acestea includ enzime digestive - chimotripsina, tripsina, elastaza si multi factori de coagulare a sangelui. Aceste proteine au aminoacizi identici în 40% din pozițiile lor și o conformație foarte asemănătoare (Fig. 1.29).

Orez. 1.29. Structuri spațiale ale elastazei (A) și chimotripsinei (B)
Unele substituții de aminoacizi au condus la modificări ale specificității substratului acestor proteine și la apariția diversității funcționale în cadrul familiei.
3. Familia imunoglobulinelor.În funcționarea sistemului imunitar, proteinele superfamiliei imunoglobulinelor joacă un rol imens, care include trei familii de proteine:
Anticorpi (imunoglobuline);
receptorii limfocitelor T;
Proteine ale complexului major de histocompatibilitate - MHC clasele 1 și 2 (Complex major de histocompatibilitate).
Toate aceste proteine au o structură de domeniu, constau din domenii asemănătoare imunologice omoloage și îndeplinesc funcții similare: interacționează cu structuri străine, fie dizolvate în sânge, limfă sau lichid intercelular (anticorpi), fie situate la suprafața celulelor (proprii sau străin).
4. Anticorpi- proteine specifice produse de limfocitele B ca răspuns la intrarea unei structuri străine în organism, numite antigen.
Caracteristicile structurii anticorpilor
Cele mai simple molecule de anticorpi constau din patru lanțuri polipeptidice: două ușoare identice - L, care conțin aproximativ 220 de aminoacizi, și două grele identice - H, constând din 440-700 de aminoacizi. Toate cele patru lanțuri din molecula de anticorp sunt conectate prin multe legături necovalente și patru legături disulfurice (Fig. 1.30).
Lanțurile ușor de anticorpi constau din două domenii: un domeniu variabil (VL), situat în regiunea N-terminală a lanțului polipeptidic și un domeniu constant (CL), situat la capătul C-terminal. Lanțurile grele au de obicei patru domenii: o variabilă (VH), situată la capătul N-terminal și trei domenii constante (CH1, CH2, CH3) (vezi Fig. 1.30). Fiecare domeniu de imunoglobulină are o suprastructură β-sheet în care două reziduuri de cisteină sunt legate printr-o legătură disulfurică.
Între cele două domenii constante CH1 și CH2 există o regiune care conține un număr mare de resturi de prolină, care împiedică formarea unei structuri secundare și interacțiunea lanțurilor H învecinate în acest segment. Această regiune balama conferă moleculei de anticorp flexibilitate. Între domeniile variabile ale lanțurilor grele și ușoare există două situsuri identice de legare la antigen (situuri active pentru legare antigene), prin urmare astfel de anticorpi sunt adesea numiți bivalente. Nu întreaga secvență de aminoacizi a regiunilor variabile ale ambelor lanțuri este implicată în legarea antigenului de anticorp, ci doar 20-30 de aminoacizi localizați în regiunile hipervariabile ale fiecărui lanț. Aceste regiuni determină capacitatea unică a fiecărui tip de anticorp de a interacționa cu antigenul complementar corespunzător.
Anticorpii sunt una dintre liniile de apărare ale organismului împotriva organismelor străine invadatoare. Funcționarea lor poate fi împărțită în două etape: prima etapă este recunoașterea și legarea antigenului pe suprafața organismelor străine, ceea ce este posibil datorită prezenței situsurilor de legare a antigenului în structura anticorpului; a doua etapă este inițierea procesului de inactivare și distrugere a antigenului. Specificitatea etapei a doua depinde de clasa de anticorpi. Există cinci clase de lanțuri grele, care diferă unele de altele în structura domeniilor constante: α, δ, ε, γ și μ, conform cărora se disting cinci clase de imunoglobuline: A, D, E, G și M.
Caracteristicile structurale ale lanțurilor grele conferă regiunilor balama și regiunilor C-terminale ale lanțurilor grele o conformație caracteristică fiecărei clase. După ce antigenul se leagă de un anticorp, modificările conformaționale în domeniile constante determină calea pentru îndepărtarea antigenului.
 Orez. 1. 30. Structura domeniului IgG
Orez. 1. 30. Structura domeniului IgG
Imunoglobulinele M
Imunoglobulinele M au două forme.
Forma monomerică- clasa I de anticorpi produși prin dezvoltarea limfocitelor B. Ulterior, multe celule B trec la producerea altor clase de anticorpi, dar cu același situs de legare a antigenului. IgM este încorporată în membrană și acționează ca un receptor de recunoaștere a antigenului. Integrarea IgM în membrana celulară este posibilă datorită prezenței a 25 de resturi de aminoacizi hidrofobe în partea de coadă a regiunii.
Forma secretorie a IgM conține cinci subunități monomerice legate între ele prin legături disulfurice și un lanț J polipeptidic suplimentar (Fig. 1.31). Lanțurile grele de monomeri de această formă nu conțin o coadă hidrofobă. Pentamerul are 10 situsuri de legare a antigenului și, prin urmare, este eficient în recunoașterea și îndepărtarea antigenului care intră mai întâi în organism. Forma secretorie a IgM este clasa principală de anticorpi secretați în sânge în timpul răspunsului imun primar. Legarea IgM la un antigen modifică conformația IgM și induce legarea acesteia la prima componentă proteică a sistemului complement (sistemul complement este un set de proteine implicate în distrugerea antigenului) și activarea acestui sistem. Dacă antigenul este situat pe suprafața unui microorganism, sistemul de complement provoacă perturbarea integrității membranei celulare și moartea celulei bacteriene.
Imunoglobulinele G
Cantitativ, această clasă de imunoglobuline predomină în sânge (75% din toate Ig-urile). IgG - monomeri, clasa principală de anticorpi secretați în sânge în timpul unui răspuns imun secundar. După interacțiunea IgG cu antigenele de suprafață ale microorganismelor, complexul antigen-anticorp este capabil să lege și să activeze proteinele sistemului complement sau poate interacționa cu receptorii specifici ai macrofagelor și neutrofilelor. Interacțiunea cu fagocitele conduce
 Orez. 1.31. Structura formei secretoare a IgM
Orez. 1.31. Structura formei secretoare a IgM
la absorbția complexelor antigen-anticorp și distrugerea lor în fagozomii celulari. IgG este singura clasă de anticorpi care sunt capabili să pătrundă în bariera placentară și să ofere protecție intrauterină a fătului împotriva infecțiilor.
Imunoglobulinele A
Clasa principală de anticorpi prezenți în secreții (lapte, saliva, secreții ale tractului respirator și tractului intestinal). IgA este secretată în principal sub formă dimerică, unde monomerii sunt legați unul de celălalt printr-un lanț J suplimentar (Fig. 1.32).
IgA nu interacționează cu sistemul complement și cu celulele fagocitare, dar prin legarea de microorganisme, anticorpii împiedică atașarea lor de celulele epiteliale și pătrunderea în organism.
Imunoglobulinele E
Imunoglobulinele E sunt reprezentate de monomeri în care lanțurile ε grele conțin, ca și lanțurile μ ale imunoglobulinelor M, un domeniu variabil și patru constante. După secreție, IgE se leagă de ea
 Orez. 1.32. Structura IgA
Orez. 1.32. Structura IgA
Regiunile C-terminale cu receptori corespunzători pe suprafața mastocitelor și bazofilelor. Ca urmare, ei devin receptori pentru antigene de pe suprafața acestor celule (Fig. 1.33).
 Orez. 1.33. Interacțiunea IgE cu antigenul de pe suprafața unui mastocit
Orez. 1.33. Interacțiunea IgE cu antigenul de pe suprafața unui mastocit
După ce antigenul se atașează la situsurile corespunzătoare de legare la antigen ale IgE, celulele primesc un semnal pentru a secreta substanțe biologic active (histamină, serotonină), care sunt în mare parte responsabile pentru dezvoltarea reacției inflamatorii și pentru manifestarea reacțiilor alergice, cum ar fi: astm, urticarie, febra fânului.
Imunoglobuline D
Imunoglobulinele D se găsesc în cantități foarte mici în ser; sunt monomeri. Lanțurile grele δ au o variabilă și trei domenii constante. IgD acționează ca receptori pentru limfocitele B; alte funcții sunt încă necunoscute. Interacțiunea antigenelor specifice cu receptorii de pe suprafața limfocitelor B (IgD) duce la transmiterea acestor semnale în celulă și la activarea mecanismelor care asigură proliferarea unei anumite clone de limfocite.
TEMA 1.7. PROPRIETĂȚI FIZICE ȘI CHIMICE ALE PROTEINELOR ȘI METODE DE SEPARARE A LOR
1. Proteinele individuale diferă în proprietăți fizice și chimice:
Forma moleculelor;
Greutate moleculară;
Sarcina totală, a cărei mărime depinde de raportul dintre grupările anionice și cationice ale aminoacizilor;
Raportul dintre radicalii de aminoacizi polari și nepolari de pe suprafața moleculelor;
Grade de rezistență la diferiți agenți de denaturare.
2. Solubilitatea proteinelor depinde asupra proprietăților proteinelor enumerate mai sus, precum și asupra compoziției mediului în care se dizolvă proteina (valori pH, compoziția sării, temperatură, prezența altor substanțe organice care pot interacționa cu proteina). Cantitatea de încărcare a moleculelor de proteine este unul dintre factorii care afectează solubilitatea acestora. Când sarcina din punctul izoelectric se pierde, proteinele se agregează și precipită mai ușor. Acest lucru este tipic în special pentru proteinele denaturate, în care radicalii de aminoacizi hidrofobi apar la suprafață.
Pe suprafața unei molecule de proteine există atât radicali de aminoacizi încărcați pozitiv, cât și negativ. Numărul acestor grupe și, prin urmare, încărcătura totală a proteinelor, depinde de pH-ul mediului, adică. raportul concentrațiilor grupelor H+ - și OH -. Într-un mediu acid O creștere a concentrației de H+ duce la suprimarea disocierii grupărilor carboxil -COO - + H+ > - COOH și la scăderea sarcinii negative a proteinelor. Într-un mediu alcalin, legarea excesului de OH - de către protonii formați în timpul disocierii grupărilor amino -NH 3 + + OH - - NH 2 + H 2 O cu formarea apei, duce la scăderea sarcinii pozitive a proteinelor. . Se numește valoarea pH-ului la care o proteină are o sarcină netă zero punct izoelectric (IEP).În IET, numărul de grupuri încărcate pozitiv și negativ este același, adică. proteina este în stare izoelectrică.
3. Separarea proteinelor individuale. Caracteristicile structurii și funcționării organismului depind de setul de proteine sintetizate în acesta. Studierea structurii și proprietăților proteinelor este imposibilă fără a le izola de celulă și a le purifica de alte proteine și molecule organice. Etape de izolare și purificare a proteinelor individuale:
Distrugerea celulelorţesutul fiind studiat şi obţinerea unui omogenat.
Separarea omogenatului în fracțiuni prin centrifugare, obținând o fracție nucleară, mitocondrială, citosolică sau altă fracție care conține proteina dorită.
Denaturarea termică selectivă- încălzirea pe termen scurt a unei soluții proteice, timp în care unele dintre impuritățile proteice denaturate pot fi îndepărtate (dacă proteina este relativ stabilă la căldură).
Sărare. Diferite proteine precipită la diferite concentrații de sare în soluție. Prin creșterea treptată a concentrației de sare, este posibilă obținerea unui număr de fracții separate cu un conținut predominant de proteine izolate într-una dintre ele. Sulfatul de amoniu este cel mai adesea folosit pentru fracţionarea proteinelor. Proteinele cu cea mai mică solubilitate precipită la concentrații scăzute de sare.
Filtrare pe gel- o metodă de cernere a moleculelor prin granule Sephadex umflate (lanțuri polizaharide tridimensionale de dextran având pori). Viteza cu care proteinele trec printr-o coloană umplută cu Sephadex va depinde de greutatea lor moleculară: cu cât masa moleculelor proteice este mai mică, cu atât ele mai ușor pătrund în granule și rămân acolo mai mult timp; cu cât masa este mai mare, cu atât eluează mai repede din coloană.
Ultracentrifugarea- o metodă care presupune introducerea proteinelor într-un tub de centrifugă în rotorul unei ultracentrifugă. Când rotorul se rotește, viteza de sedimentare a proteinelor este proporțională cu greutatea moleculară a acestora: fracțiile de proteine mai grele sunt situate mai aproape de fundul eprubetei, cele mai ușoare - mai aproape de suprafață.
Electroforeză- o metodă bazată pe diferențele de viteză de mișcare a proteinelor într-un câmp electric. Această valoare este proporțională cu sarcina proteinelor. Electroforeza proteinelor se efectuează pe hârtie (în acest caz, viteza de mișcare a proteinelor este proporțională doar cu sarcina lor) sau într-un gel de poliacrilamidă cu o anumită dimensiune a porilor (viteza de mișcare a proteinelor este proporțională cu sarcina și greutatea moleculară a acestora) .
Cromatografia cu schimb de ioni- o metodă de fracţionare bazată pe legarea grupărilor ionizate de proteine cu grupări încărcate opus de răşini schimbătoare de ioni (materiale polimerice insolubile). Puterea legării proteinei de rășină este proporțională cu sarcina proteinei. Proteinele adsorbite la polimerul schimbător de ioni pot fi spălate cu concentrații crescânde de soluții de NaCl; cu cât este mai mică sarcina de proteine, cu atât este mai mică concentrația de NaCl necesară pentru a spăla proteina legată de grupările ionice ale rășinii.
Cromatografia de afinitate- metoda cea mai specifica de izolare a proteinelor individuale.Un ligand al unei proteine este atasat covalent de un polimer inert. Când o soluție de proteină este trecută printr-o coloană cu un polimer, numai proteina specifică pentru un ligand dat este adsorbită pe coloană datorită legării complementare a proteinei la ligand.
Dializă- o metodă folosită pentru îndepărtarea compușilor cu greutate moleculară mică dintr-o soluție de proteină izolată. Metoda se bazează pe incapacitatea proteinelor de a trece printr-o membrană semi-permeabilă, spre deosebire de substanțele cu greutate moleculară mică. Este folosit pentru a purifica proteinele din impuritățile cu molecularitate scăzută, de exemplu, sărurile după sărare.
TECTURI PENTRU MUNCĂ EXTRAȘCOLARĂ
1. Completați tabelul. 1.4.
Tabelul 1.4. Analiza comparativă a structurii și funcțiilor proteinelor înrudite - mioglobina și hemoglobina
a) amintiți-vă structura centrului activ al lui Mb și Hb. Ce rol joacă radicalii de aminoacizi hidrofobi în formarea centrilor activi ai acestor proteine? Descrieți structura centrului activ al Mb și Hb și mecanismele de atașare a liganzilor la acesta. Ce rol joacă reziduurile His F 8 și His E 7 în funcționarea centrului activ al Mv iHv?
b) ce proprietăți noi are în comparație cu mioglobina monomerică proteina oligomerică strâns înrudită, hemoglobina? Explicați rolul modificărilor cooperante în conformația protomerilor din molecula de hemoglobină, efectul concentrațiilor de CO 2 și protoni asupra afinității hemoglobinei pentru oxigen, precum și rolul 2,3-BPG în reglarea alosterică a funcției Hb .
2. Caracterizați însoțitorii moleculari, acordând atenție relației dintre structura și funcția lor.
3. Ce proteine sunt grupate în familii? Folosind exemplul familiei de imunoglobuline, identificați caracteristicile structurale similare și funcțiile înrudite ale proteinelor acestei familii.
4. Proteinele individuale purificate sunt adesea necesare în scopuri biochimice și medicinale. Explicați pe ce proprietăți fizico-chimice se bazează metodele utilizate pentru separarea și purificarea proteinelor.
SARCINI DE AUTOCONTROL
1. Alege raspunsurile corecte.
Funcțiile hemoglobinei:
A. Transportul O 2 de la plămâni la țesut B. Transportul de H + de la țesuturi la plămâni
B. Menținerea unui pH constant al sângelui D. Transportul CO 2 de la plămâni la țesuturi
D. Transportul CO 2 din țesuturi la plămâni
2. Alege raspunsurile corecte. Ligandα -protomerul Hb este: A. Heme
B. Oxigen
B. CO G. 2,3-BPG
D. β-Protomer
3. Alege raspunsurile corecte.
Hemoglobina spre deosebire de mioglobina:
A. Are o structură cuaternară
B. Structura secundară este reprezentată doar de elice α
B. Aparține proteinelor complexe
D. Interacționează cu un ligand alosteric D. Legat covalent de hem
4. Alege raspunsurile corecte.
Afinitatea Hb pentru O2 scade:
A. Când se adaugă o moleculă de O 2 B. Când o moleculă de O 2 este îndepărtată
B. Când interacționați cu 2,3-BPG
D. Când este atașat de protomeri H + D. Când concentrația de 2,3-BPG scade
5. Meci.
Tipurile HB se caracterizează prin:
A. În formă deoxi formează agregate fibrilare B. Conține două lanțuri α și două δ
B. Forma predominantă de Hb în eritrocitele adulte D. Conține hem cu Fe+ 3 în centrul activ
D. Conține două lanțuri α și două γ 1. HbA 2.
6. Meci.
Liganzii Hb:
A. Se leagă de Hb în centrul alosteric
B. Are o afinitate foarte mare pentru situsul activ al Hb
B. Prin unire, crește afinitatea Hb pentru O 2 G. Oxidează Fe+ 2 la Fe+ 3
D. Formează o legătură covalentă cu hisF8
7. Alege raspunsurile corecte.
Însoțitori:
A. Proteine prezente în toate părțile celulei
B. Sinteza crește sub stres
B. Participa la hidroliza proteinelor denaturate
D. Participa la menținerea conformației native a proteinelor
D. Ele creează organite în care se formează conformația proteinelor.
8. Meci. Imunoglobuline:
A. Forma secretorie este pentamerică.
B. Clasa Ig care pătrunde în bariera placentară
B. Ig - receptor mastocitar
D. Clasa principală de Ig prezentă în secrețiile celulelor epiteliale. D. Receptorul limfocitelor B, a cărui activare asigură proliferarea celulară
9. Alege raspunsurile corecte.
Imunoglobulinele E:
A. Produse de macrofage B. Au lanțuri ε grele.
B. Înglobat în membrana limfocitelor T
D. Acționează ca receptori de antigen de membrană pe mastocite și bazofile
D. Responsabil pentru reacțiile alergice
10. Alege raspunsurile corecte.
Metoda de separare a proteinelor se bazează pe diferențele de greutate moleculară a acestora:
A. Filtrare pe gel
B. Ultracentrifugarea
B. Electroforeza pe gel de poliacrilamidă D. Cromatografia de schimb ionic
D. Cromatografia de afinitate
11. Alege răspunsul corect.
Metoda de separare a proteinelor se bazează pe diferențele de solubilitate a acestora în apă:
A. Filtrare pe gel B. Sărare
B. Cromatografia de schimb ionic D. Cromatografia de afinitate
D. Electroforeza în gel de poliacrilamidă
STANDARDE DE RĂSPUNSURI LA „SARCINI DE AUTOCONTROL”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
TERMENI ȘI CONCEPTE DE BAZĂ
1. Proteine oligomerice, protomer, structura cuaternară a proteinelor
2. Modificări cooperante în conformația protomerului
3. Efectul Bohr
4. Reglarea alosterică a funcțiilor proteice, centru alosteric și efector alosteric
5. Chaperone moleculare, proteine de șoc termic
6. Familii de proteine (serin proteaze, imunoglobuline)
7. Relația IgM-, G-, E-, A-structură-funcție
8. Sarcina totală a proteinelor, punctul izoelectric al proteinelor
9. Electroforeza
10. Sărare
11. Filtrare pe gel
12. Cromatografia de schimb ionic
13. Ultracentrifugarea
14. Cromatografia de afinitate
15. Electroforeza proteinelor plasmatice sanguine
SARCINI PENTRU MUNCĂ LA CLASĂ
1. Comparați dependențele gradelor de saturație ale hemoglobinei (Hb) și mioglobinei (Mb) cu oxigenul de presiunea sa parțială în țesuturi
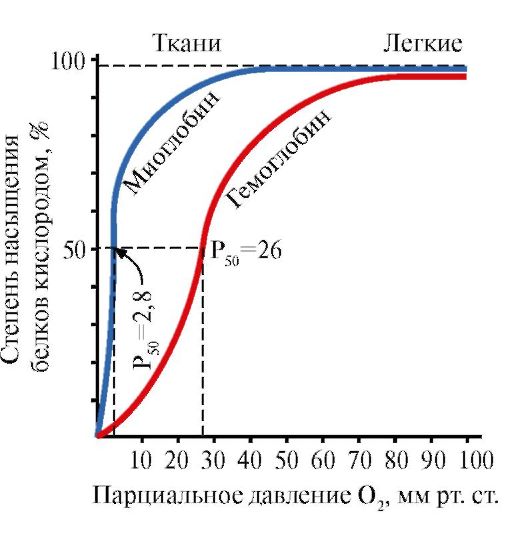 Orez. 1.34. Dependenţa de saturaţie Mv şiNHoxigen din presiunea sa parțială
Orez. 1.34. Dependenţa de saturaţie Mv şiNHoxigen din presiunea sa parțială
Vă rugăm să rețineți că forma curbelor de saturație a oxigenului proteinei este diferită: pentru mioglobină - o hiperbolă, pentru hemoglobină - o formă sigmoidă.
1. comparați valorile presiunii parțiale a oxigenului la care Mb și Hb sunt saturate cu O 2 cu 50%. Care dintre aceste proteine are o afinitate mai mare pentru O2?
2. Ce caracteristici structurale ale Mb determină afinitatea sa mare pentru O 2?
3. Ce caracteristici structurale ale HB îi permit să elibereze O2 în capilarele țesuturilor în repaus (la o presiune parțială relativ mare a O2) și să mărească brusc această eliberare în mușchii care lucrează? Ce proprietate a proteinelor oligomerice oferă acest efect?
4. Calculați ce cantitate de O 2 (în %) o dă hemoglobina oxigenată mușchilor care se odihnesc și lucrează?
5. trageți concluzii despre relația dintre structura unei proteine și funcția acesteia.
2. Cantitatea de oxigen eliberată de hemoglobină în capilare depinde de intensitatea proceselor catabolice din țesuturi (efect Bohr). Cum modificările metabolismului tisular reglează afinitatea Hb pentru O2? Efectul CO2 și H+ asupra afinității Hb pentru O2
 1. descrie efectul Bohr.
1. descrie efectul Bohr.
2. în ce direcție se desfășoară procesul prezentat în diagramă:
a) în capilarele plămânilor;
b) în capilarele tisulare?
3. Care este semnificația fiziologică a efectului Bohr?
4. De ce interacțiunea Hb cu H+ la locuri îndepărtate de hem modifică afinitatea proteinei pentru O 2?
3. Afinitatea Hb pentru O2 depinde de concentrația ligandului său - 2,3-bisfosfoglicerat, care este un regulator alosteric al afinității Hb pentru O2. De ce interacțiunea ligandului la un loc îndepărtat de locul activ afectează funcția proteinei? Cum reglează 2,3-BPG afinitatea Hb pentru O2? Pentru a rezolva problema, răspunde la următoarele întrebări:
1. unde și din ce se sintetizează 2,3-bisfosfogliceratul (2,3-BPG)? Scrieți formula acesteia, indicați sarcina acestei molecule.
2. Cu ce formă de hemoglobină (oxi sau deoxi) interacționează BPG și de ce? În ce parte a moleculei de Hb are loc interacțiunea?
3. în ce direcție are loc procesul prezentat în diagramă?
a) în capilarele tisulare;
b) în capilarele plămânilor?
4. unde concentraţia complexului ar trebui să fie mai mare
Nv-2,3-BFG:
a) în capilarele mușchilor în repaus,
b) în capilarele mușchilor care lucrează (cu condiția aceeași concentrație de BPG în eritrocite)?
5. Cum se va schimba afinitatea HB pentru oxigen atunci când o persoană se adaptează la condițiile de altitudine mare, dacă crește concentrația de BPG în eritrocite? Care este semnificația fiziologică a acestui fenomen?
4. Distrugerea 2,3-BPG în timpul depozitării sângelui conservat afectează funcțiile HB. Cum se va schimba afinitatea HB pentru O 2 în sângele conservat dacă concentrația de 2,3-BPG în eritrocite poate scădea de la 8 la 0,5 mmol/l. Este posibil să se transfuzeze un astfel de sânge pacienților grav bolnavi dacă concentrația de 2,3-BPG este restabilită nu mai devreme de trei zile? Este posibil să restabiliți funcțiile celulelor roșii din sânge prin adăugarea de 2,3-BPG în sânge?
5. Amintiți-vă structura celor mai simple molecule de imunoglobuline. Ce rol joacă imunoglobulinele în sistemul imunitar? De ce Ig-urile sunt adesea numite bivalente? Cum se raportează structura Ig-urilor cu funcția lor? (Descrieți folosind un exemplu de clasă de imunoglobuline.)
Proprietățile fizico-chimice ale proteinelor și metodele de separare a acestora.
6. Cum afectează încărcătura netă a unei proteine solubilitatea acesteia?
a) determinați sarcina totală a peptidei la pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) cum se va schimba sarcina acestei peptide la pH >7, pH<7, рН <<7?
c) care este punctul izoelectric al unei proteine (IEP) și în ce mediu se află?
IET al acestei peptide?
d) la ce valoare pH se va observa cea mai mică solubilitate a acestei peptide.
7. De ce laptele acru, spre deosebire de laptele proaspăt, se „coagulează” atunci când este fiert (adică precipită cazeina din proteinele din lapte)? În laptele proaspăt, moleculele de cazeină au o sarcină negativă.
8. Filtrarea pe gel este utilizată pentru a separa proteinele individuale. Un amestec care conține proteine A, B, C cu greutăți moleculare egale cu 160.000, 80.000 și, respectiv, 60.000, a fost analizat prin filtrare pe gel (Fig. 1.35). Granulele de gel umflate sunt permeabile la proteinele cu o greutate moleculară mai mică de 70 000. Ce principiu stă la baza acestei metode de separare? Care grafic reflectă corect rezultatele fracționării? Indicați ordinea în care proteinele A, B și C sunt eliberate din coloană.
 Orez. 1.35. Utilizarea gelului de filtrare pentru separarea proteinelor
Orez. 1.35. Utilizarea gelului de filtrare pentru separarea proteinelor
9. În fig. 1.36, A prezintă o diagramă a electroforezei pe hârtie a proteinelor din serul sanguin al unei persoane sănătoase. Cantitățile relative de fracții proteice obținute prin această metodă sunt: albumine 54-58%, α1-globuline 6-7%, α2-globuline 8-9%, β-globuline 13%, y-globuline 11-12%.
 Orez. 1.36 Electroforeza pe hârtie a proteinelor plasmatice ale unei persoane sănătoase (A) și ale unui pacient (B)
Orez. 1.36 Electroforeza pe hârtie a proteinelor plasmatice ale unei persoane sănătoase (A) și ale unui pacient (B)
I - y-globuline; II - β-globuline; III -α 2 -globulina; IV -α 2 -globulina; V - albumine
Multe boli sunt însoțite de modificări cantitative ale compoziției proteinelor serice (disproteinemie). Natura acestor modificări este luată în considerare atunci când se pune un diagnostic și se evaluează severitatea și stadiul bolii.
Folosind datele din tabel. 1.5, faceți o ghicire despre boală, care se caracterizează prin profilul electroforetic prezentat în Fig. 1.36.
Tabelul 1.5. Modificări ale concentrației proteinelor serice în patologie
S-a dovedit existența a 4 niveluri de organizare structurală a unei molecule proteice.
Structura primară a proteinei– secvența de aranjare a resturilor de aminoacizi în lanțul polipeptidic. În proteine, aminoacizii individuali sunt legați unul de celălalt legături peptidice, care rezultă din interacțiunea grupărilor a-carboxil și a-amino ale aminoacizilor.
Până în prezent, structura primară a zeci de mii de proteine diferite a fost descifrată. Pentru a determina structura primară a unei proteine, compoziția de aminoacizi este determinată folosind metode de hidroliză. Apoi se determină natura chimică a aminoacizilor terminali. Următorul pas este determinarea secvenței de aminoacizi din lanțul polipeptidic. În acest scop, se utilizează hidroliza parțială selectivă (chimică și enzimatică). Este posibil să se utilizeze analiza de difracție cu raze X, precum și date despre secvența de nucleotide complementară a ADN-ului.
Structura secundară a proteinei– configurația lanțului polipeptidic, adică o metodă de împachetare a unui lanț polipeptidic într-o conformație specifică. Acest proces nu decurge haotic, ci în conformitate cu programul încorporat în structura primară.
Stabilitatea structurii secundare este asigurată în principal de legăturile de hidrogen, dar o anumită contribuție o au legăturile covalente - peptidă și disulfură.
Este considerat cel mai probabil tip de structură a proteinelor globulare a-helix. Răsucirea lanțului polipeptidic are loc în sensul acelor de ceasornic. Fiecare proteină este caracterizată printr-un anumit grad de elicoidalizare. Dacă lanțurile hemoglobinei sunt 75% elicoidale, atunci pepsina este de doar 30%.
Se numește tipul de configurație a lanțurilor polipeptidice găsite în proteinele părului, mătăsii și mușchilor b-structuri. Segmentele lanțului peptidic sunt aranjate într-un singur strat, formând o figură asemănătoare unei foi pliate într-un acordeon. Stratul poate fi format din două sau mai multe lanțuri peptidice.
În natură, există proteine a căror structură nu corespunde nici structurii β sau a, de exemplu, colagenul este o proteină fibrilă care alcătuiește cea mai mare parte a țesutului conjunctiv din corpul uman și animal.
Structura terțiară a proteinelor– orientarea spațială a helixului polipeptidic sau modul în care lanțul polipeptidic este așezat într-un anumit volum. Prima proteină a cărei structură terțiară a fost elucidată prin analiza de difracție cu raze X a fost mioglobina de cașlot (Fig. 2).
În stabilizarea structurii spațiale a proteinelor, pe lângă legăturile covalente, rolul principal îl au legăturile necovalente (hidrogen, interacțiuni electrostatice ale grupărilor încărcate, forțe intermoleculare van der Waals, interacțiuni hidrofobe etc.).
Conform conceptelor moderne, structura terțiară a unei proteine, după terminarea sintezei sale, se formează spontan. Forța motrice principală este interacțiunea radicalilor de aminoacizi cu moleculele de apă. În acest caz, radicalii de aminoacizi hidrofobi nepolari sunt scufundați în interiorul moleculei de proteine, iar radicalii polari sunt orientați spre apă. Procesul de formare a structurii spațiale native a unui lanț polipeptidic este numit pliere. Proteine numite însoţitori. Ei participă la pliere. Au fost descrise o serie de boli ereditare umane, a căror dezvoltare este asociată cu tulburări datorate mutațiilor în procesul de pliere (pigmentoză, fibroză etc.).
Prin metodele de analiză prin difracție de raze X s-a dovedit existența unor niveluri de organizare structurală a moleculei proteice, intermediare între structurile secundare și terțiare. Domeniu este o unitate structurală globulară compactă în cadrul unui lanț polipeptidic (Fig. 3). Au fost descoperite multe proteine (de exemplu, imunoglobuline), constând din domenii de structură și funcții diferite, codificate de gene diferite.
Toate proprietățile biologice ale proteinelor sunt asociate cu păstrarea structurii lor terțiare, care se numește nativ. Globulul proteic nu este o structură absolut rigidă: sunt posibile mișcări reversibile ale unor părți ale lanțului peptidic. Aceste modificări nu perturbă conformația generală a moleculei. Conformația unei molecule de proteine este influențată de pH-ul mediului, puterea ionică a soluției și interacțiunea cu alte substanțe. Orice influență care duce la perturbarea conformației native a moleculei este însoțită de pierderea parțială sau completă a proprietăților biologice ale proteinei.
Structura proteinelor cuaternare- o metodă de așezare în spațiu a lanțurilor polipeptidice individuale care au aceeași structură primară, secundară sau terțiară aceeași sau diferită și formarea unei formațiuni macromoleculare unificate structural și funcțional.
Se numește o moleculă de proteină constând din mai multe lanțuri polipeptidice oligomerși fiecare lanț inclus în el - protomer. Proteinele oligomerice sunt adesea construite dintr-un număr par de protomeri, de exemplu, molecula de hemoglobină constă din două lanțuri polipeptidice a și două b (Fig. 4).
Aproximativ 5% dintre proteine au o structură cuaternară, incluzând hemoglobina și imunoglobulinele. Structura subunității este caracteristică multor enzime.
Moleculele proteice care alcătuiesc o proteină cu structură cuaternară se formează separat pe ribozomi și numai după terminarea sintezei formează o structură supramoleculară comună. O proteină dobândește activitate biologică numai atunci când protomerii ei constituenți sunt combinați. La stabilizarea structurii cuaternare participă aceleași tipuri de interacțiuni ca și la stabilizarea structurii terțiare.
Unii cercetători recunosc existența unui al cincilea nivel de organizare structurală a proteinelor. Acest metabolii - complexe macromoleculare polifuncționale de diverse enzime care catalizează întreaga cale de transformări a substratului (sintetaze superioare de acizi grași, complex de piruvat dehidrogenază, lanț respirator).
Structura modulului | Teme |
Unitate modulară 1 | 1.1. Organizarea structurală a proteinelor. Etape de formare a conformației native a proteinelor 1.2. Bazele funcționării proteinelor. Medicamentele ca liganzi care afectează funcția proteinelor 1.3. Denaturarea proteinelor și posibilitatea renativării lor spontane |
Unitatea modulară 2 | 1.4. Caracteristici ale structurii și funcționării proteinelor oligomerice folosind exemplul hemoglobinei 1.5. Menținerea conformației proteinelor native în condiții celulare 1.6. Varietate de proteine. Familii de proteine folosind exemplul imunoglobulinelor 1.7. Proprietățile fizico-chimice ale proteinelor și metodele de separare a acestora |
Unitatea modulara 1 ORGANIZAREA STRUCTURALA A PROTEINELOR MONOMERICE SI BAZELE FUNCTIONARII LOR
Obiective de învățare A fi capabil să:
1. Utilizați cunoștințele despre caracteristicile structurale ale proteinelor și dependența funcțiilor proteinelor de structura lor pentru a înțelege mecanismele de dezvoltare a proteinopatiilor ereditare și dobândite.
2. Explicați mecanismele acțiunii terapeutice a unor medicamente ca liganzi care interacționează cu proteinele și își modifică activitatea.
3. Utilizați cunoștințele despre structura și labilitatea conformațională a proteinelor pentru a înțelege instabilitatea lor structurală și funcțională și tendința de denaturare în condiții în schimbare.
4. Explicați utilizarea agenților de denaturare ca mijloace de sterilizare a materialelor și instrumentelor medicale, precum și a antisepticelor.
Știi:
1. Niveluri de organizare structurală a proteinelor.
2. Importanța structurii primare a proteinelor, care determină diversitatea lor structurală și funcțională.
3. Mecanismul de formare a centrului activ în proteine și interacțiunea specifică a acestuia cu ligand, care stă la baza funcționării proteinelor.
4. Exemple de influență a liganzilor exogeni (medicamente, toxine, otrăvuri) asupra conformației și activității funcționale a proteinelor.
5. Cauze si consecinte ale denaturarii proteinelor, factori care determina denaturarea.
6. Exemple de utilizare a factorilor denaturanți în medicină ca antiseptice și mijloace pentru sterilizarea instrumentelor medicale.
TEMA 1.1. ORGANIZAREA STRUCTURALĂ A PROTEINELOR. ETAPE DE FORMARE A NATIVULUI
CONFORMATII PROTEINE
Proteinele sunt molecule polimerice ai căror monomeri sunt doar 20 de α-aminoacizi. Setul și ordinea combinației de aminoacizi într-o proteină este determinată de structura genelor din ADN-ul indivizilor. Fiecare proteină, în conformitate cu structura sa specifică, își îndeplinește propria funcție. Setul de proteine ale unui anumit organism determină caracteristicile sale fenotipice, precum și prezența bolilor ereditare sau o predispoziție la dezvoltarea lor.
1. Aminoacizi care alcătuiesc proteinele. Legătură peptidică. Proteinele sunt polimeri formați din monomeri - 20 de α-aminoacizi, a căror formulă generală este
Aminoacizii diferă în ceea ce privește structura, dimensiunea și proprietățile fizico-chimice ale radicalilor atașați la atomul de carbon α. Grupurile funcționale ale aminoacizilor determină caracteristicile proprietăților diferiților α-aminoacizi. Radicalii găsiți în α-aminoacizi pot fi împărțiți în mai multe grupuri:
prolina, Spre deosebire de ceilalți 19 monomeri proteici, nu este un aminoacid, ci un iminoacid; radicalul din prolină este asociat atât cu atomul de carbon α, cât și cu gruparea imino.
Aminoacizii variază în solubilitate în apă. Acest lucru se datorează capacității radicalilor de a interacționa cu apa (hidrat).
LA hidrofil includ radicali care conțin grupări funcționale neîncărcate anionice, cationice și polare.
LA hidrofob includ radicali care conțin grupări metil, lanțuri alifatice sau inele.
2. Legăturile peptidice conectează aminoacizii pentru a forma peptide.În timpul sintezei peptidelor, gruparea α-carboxil a unui aminoacid interacționează cu gruparea α-amino a altui aminoacid pentru a forma legătură peptidică:
Proteinele sunt polipeptide, adică. polimeri liniari ai α-aminoacizilor legați printr-o legătură peptidică (Fig. 1.1.)
Orez. 1.1. Termeni folosiți pentru a descrie structura peptidelor
Monomerii aminoacizilor care formează polipeptidele se numesc reziduuri de aminoacizi. Un lanț de grupuri care se repetă - NH-CH-CO- forme coloana vertebrală peptidică. Un rest de aminoacid care are o grupare α-amino liberă se numește N-terminal, iar unul care are o grupare α-carboxil liberă se numește C-terminal. Peptidele sunt scrise și citite de la capătul N-terminal la capătul C-terminal.
Legătura peptidică formată de grupul imino a prolinei diferă de alte legături peptidice: atomul de azot al grupului peptidic nu are hidrogen,
în schimb, există o legătură cu un radical, ca urmare a căreia o parte a inelului este inclusă în scheletul peptidic:
Peptidele diferă în ceea ce privește compoziția de aminoacizi, numărul de aminoacizi și ordinea conexiunii aminoacizilor, de exemplu, Ser-Ala-Glu-Gis și His-Glu-Ala-Ser sunt două peptide diferite.
Legăturile peptidice sunt foarte puternice, iar hidroliza lor chimică neenzimatică necesită condiții dure: proteina analizată este hidrolizată în acid clorhidric concentrat la o temperatură de aproximativ 110° timp de 24 de ore. Într-o celulă vie, legăturile peptidice pot fi rupte enzime proteolitice, numit proteaze sau hidrolaze peptidice.
3. Structura primară a proteinelor. Reziduurile de aminoacizi din lanțurile peptidice ale diferitelor proteine nu alternează aleatoriu, ci sunt aranjate într-o anumită ordine. Se numește secvența liniară sau ordinea de alternanță a resturilor de aminoacizi dintr-un lanț polipeptidic structura primară a proteinei.
Structura primară a fiecărei proteine individuale este codificată în molecula de ADN (într-o regiune numită genă) și este realizată în timpul transcripției (copierea informațiilor pe ARNm) și translației (sinteza structurii primare a proteinei). În consecință, structura primară a proteinelor unei persoane individuale este informația transmisă ereditar de la părinți la copii, care determină caracteristicile structurale ale proteinelor unui anumit organism, de care depinde funcția proteinelor existente (Fig. 1.2.).
Orez. 1.2. Relația dintre genotip și conformația proteinelor sintetizate în corpul individului
Fiecare dintre cele aproximativ 100.000 de proteine individuale din corpul uman are unic structura primara. Moleculele de același tip de proteină (de exemplu, albumina) au aceeași alternanță de resturi de aminoacizi, ceea ce distinge albumina de orice altă proteină individuală.
Secvența de resturi de aminoacizi dintr-un lanț peptidic poate fi considerată o formă de înregistrare a informațiilor. Aceste informații determină aranjarea spațială a lanțului peptidic linear într-o structură tridimensională mai compactă numită conformaţie veveriţă. Procesul de formare a unei conformații proteice active funcțional se numește pliere
4. Conformația proteinei. Rotația liberă în scheletul peptidic este posibilă între atomul de azot al grupării peptidice și atomul de carbon α vecin, precum și între atomul de carbon α și carbonul grupării carbonil. Datorită interacțiunii grupurilor funcționale de reziduuri de aminoacizi, structura primară a proteinelor poate dobândi structuri spațiale mai complexe. În proteinele globulare, există două niveluri principale de pliere a conformației lanțurilor peptidice: secundarȘi structura tertiara.
Structura secundară a proteinelor este o structură spațială formată ca urmare a formării legăturilor de hidrogen între grupările funcționale -C=O și -NH- ale scheletului peptidic. În acest caz, lanțul peptidic poate dobândi structuri regulate de două tipuri: α-helicesȘi β-structuri.
ÎN α-helices se formează legături de hidrogen între atomul de oxigen al grupării carbonil și hidrogenul azotului amidic al celui de-al 4-lea aminoacid din acesta; lanțurile laterale de reziduuri de aminoacizi
sunt situate de-a lungul periferiei spiralei, fără a participa la formarea structurii secundare (Fig. 1.3.).
Radicalii în vrac, sau radicalii care poartă sarcini egale, împiedică formarea unui α-helix. Reziduul de prolină, care are o structură de inel, întrerupe α-helix, deoarece din cauza lipsei de hidrogen la atomul de azot din lanțul peptidic este imposibil să se formeze o legătură de hidrogen. Legătura dintre azot și atomul de carbon α face parte din inelul de prolină, astfel încât coloana vertebrală a peptidei devine îndoită în acest punct.
β-Structură se formează între regiunile liniare ale scheletului peptidic al unui lanț polipeptidic, formând astfel structuri pliate. Se pot forma lanțuri polipeptidice sau părți ale acestora paralel sau β-structuri antiparalele.În primul caz, terminalele N- și C-terminale ale lanțurilor peptidice care interacționează coincid, iar în al doilea au direcția opusă (Fig. 1.4).
Orez. 1.3. Structura secundară a proteinei - α-helix
Orez. 1.4. Structuri β-sheet paralele și antiparalele
Structurile β sunt indicate prin săgeți largi: A - Structura β antiparalelă. B - Structuri paralele β-sheet
În unele proteine, structurile β pot fi formate datorită formării legăturilor de hidrogen între atomii scheletului peptidic al diferitelor lanțuri polipeptidice.
Se găsește și în proteine zone cu secundar neregulat structura, care include îndoituri, bucle și întoarceri ale coloanei vertebrale polipeptidice. Ele sunt adesea localizate în locuri în care direcția lanțului peptidic se schimbă, de exemplu, atunci când se formează o structură paralelă a foii β.
Pe baza prezenței elicelor α și structurilor β, proteinele globulare pot fi împărțite în patru categorii.
Orez. 1.5. Structura secundară a mioglobinei (A) și a lanțului β a hemoglobinei (B), care conține opt elice α
Orez. 1.6. Structura secundară a domeniului triozofosfat izomerazei și piruvat kinazei
Orez. 1.7. Structura secundară a domeniului constant al imunoglobulinei (A) și a enzimei superoxid dismutază (B)
ÎN a patra categorie au inclus proteine care conțin o cantitate mică de structuri secundare regulate. Aceste proteine includ proteine mici sau metaloproteine bogate în cisteină.
Structura terțiară a proteinelor- un tip de conformatie format ca urmare a interactiunilor dintre radicalii de aminoacizi, care pot fi situati la o distanta considerabila unul de altul in lantul peptidic. Majoritatea proteinelor formează o structură spațială asemănătoare unui glob (proteine globulare).
Deoarece radicalii hidrofobi de aminoacizi tind să se combine prin așa-numitele interacțiuni hidrofobeși forțele intermoleculare van der Waals, în interiorul globului proteic se formează un miez hidrofob dens. Radicalii hidrofili ionizați și neionizați sunt localizați în principal pe suprafața proteinei și determină solubilitatea acesteia în apă.
Orez. 1.8. Tipuri de legături care apar între radicalii de aminoacizi în timpul formării structurii terțiare a unei proteine
1 - legătură ionică- apare între grupele funcționale încărcate pozitiv și negativ;
2 - legătură de hidrogen- apare intre o grupare hidrofila neincarcata si orice alta grupare hidrofila;
3 - interacțiuni hidrofobe- apar între radicalii hidrofobi;
4 - legătură disulfurică- formată din cauza oxidării grupurilor SH ale reziduurilor de cisteină și a interacțiunii lor între ele
Reziduurile de aminoacizi hidrofile situate în interiorul miezului hidrofob pot interacționa între ele folosind ionicȘi legături de hidrogen(Fig. 1.8).
Legăturile ionice și de hidrogen, precum și interacțiunile hidrofobe, sunt slabe: energia lor nu este cu mult mai mare decât energia mișcării termice a moleculelor la temperatura camerei. Conformația proteinei este menținută prin formarea multor astfel de legături slabe. Deoarece atomii care alcătuiesc o proteină sunt în mișcare constantă, este posibil să se rupă unele legături slabe și să se formeze altele, ceea ce duce la mișcări ușoare ale secțiunilor individuale ale lanțului polipeptidic. Această proprietate a proteinelor de a modifica conformația ca urmare a ruperii unora și a formării altor legături slabe se numește labilitate conformaţională.
Corpul uman are sisteme care susțin homeostaziei- constanța mediului intern în anumite limite acceptabile pentru un organism sănătos. În condiții de homeostazie, mici modificări ale conformației nu perturbă structura și funcția generală a proteinelor. Conformația activă funcțional a unei proteine se numește conformatie nativa. Modificările din mediul intern (de exemplu, concentrația de glucoză, ioni de Ca, protoni etc.) duc la modificări ale conformației și perturbarea funcțiilor proteinelor.
Structura terțiară a unor proteine este stabilizată legături disulfurice, formată ca urmare a interacțiunii grupărilor -SH a două resturi
Orez. 1.9. Formarea unei legături disulfurice într-o moleculă de proteină
cisteină (Fig. 1.9). Majoritatea proteinelor intracelulare nu au legături disulfurice covalente în structura lor terțiară. Prezența lor este caracteristică proteinelor secretate de celulă, ceea ce le asigură o mai mare stabilitate în condiții extracelulare. Astfel, legăturile disulfurice sunt prezente în moleculele de insulină și imunoglobuline.
Insulină- un hormon proteic sintetizat în celulele β ale pancreasului și secretat în sânge ca răspuns la creșterea concentrației de glucoză în sânge. În structura insulinei, există două legături disulfurice care conectează lanțurile polipeptidei A și B și o legătură disulfură în cadrul lanțului A (Fig. 1.10).
Orez. 1.10. Legături disulfurice în structura insulinei
5. Structura suprasecundară a proteinelor.În proteinele cu structură și funcții primare diferite, acestea sunt uneori detectate combinații similare și poziții relative ale structurilor secundare, care se numesc structura supersecundara. Ocupă o poziție intermediară între structurile secundare și terțiare, deoarece este o combinație specifică de elemente ale structurii secundare în formarea structurii terțiare a proteinei. Structurile supersecundare au denumiri specifice, cum ar fi „α-helix-turn-a-helix”, „fermoar leucină”, „degete de zinc” etc. Astfel de structuri supersecundare sunt caracteristice proteinelor care leagă ADN-ul.
— Fermoar leucină. Acest tip de structură supersecundară este utilizat pentru a uni două proteine împreună. Pe suprafața proteinelor care interacționează există regiuni elicoidale α care conțin cel puțin patru resturi de leucină. Reziduurile de leucină din α-helix sunt localizate la șase aminoacizi una de cealaltă. Deoarece fiecare rotație a helixului α conține 3,6 reziduuri de aminoacizi, radicalii de leucină sunt localizați pe suprafața fiecărei rotații a doua. Resturile de leucină ale α-helixului unei proteine pot interacționa cu resturile de leucină ale altei proteine (interacțiuni hidrofobe), conectându-le între ele (Fig. 1.11.). Multe proteine care leagă ADN-ul funcționează în complexe oligomerice în care subunitățile individuale sunt legate între ele prin „fermoare cu leucină”.
Orez. 1.11. „Fermoar leucină” între regiunile α-helicoidale ale două proteine
Un exemplu de astfel de proteine sunt histonele. Histones- proteine nucleare, care contin un numar mare de aminoacizi incarcati pozitiv - arginina si lizina (pana la 80%). Moleculele de histonă sunt combinate în complexe oligomerice care conțin opt monomeri folosind „fermoare de leucină”, în ciuda încărcăturii omonime semnificative a acestor molecule.
"Degetul de zinc"- o variantă de structură supersecundară, caracteristică proteinelor de legare a ADN-ului, are forma unui fragment alungit la suprafața proteinei și conține aproximativ 20 de resturi de aminoacizi (Fig. 1.12). Forma „degetului extins” este susținută de un atom de zinc legat de patru radicali de aminoacizi - două reziduuri de cisteină și două reziduuri de histidină. În unele cazuri, în loc de reziduuri de histidină, există reziduuri de cisteină. Două reziduuri de cisteină apropiate sunt separate de celelalte două resturi Gisili printr-o secvență Cys constând din aproximativ 12 resturi de aminoacizi. Această regiune a proteinei formează un α-helix, ai cărui radicali se pot lega în mod specific la regiunile reglatoare ale canelurii majore a ADN-ului. Specificitatea de legare individuală
Orez. 1.12. Structura primară a regiunii proteinelor care leagă ADN-ul care formează structura „deget de zinc” (literele indică aminoacizii care alcătuiesc această structură)
Proteina reglatoare care leagă ADN-ul depinde de secvența reziduurilor de aminoacizi situate în regiunea degetului de zinc. Astfel de structuri conțin, în special, receptori pentru hormonii steroizi implicați în reglarea transcripției (citirea informațiilor de la ADN la ARN).
TEMA 1.2. BAZELE FUNCȚIONĂRII PROTEINELOR. MEDICAMENTE CA LIGANDI AFECTAT FUNCȚIA PROTEINĂ
1. Centrul activ al proteinei și interacțiunea acestuia cu ligand.În timpul formării structurii terțiare, se formează o regiune pe suprafața unei proteine funcțional active, de obicei într-o adâncitură, formată din radicali de aminoacizi care sunt departe unul de celălalt în structura primară. Această regiune, care are o structură unică pentru o anumită proteină și este capabilă să interacționeze în mod specific cu o anumită moleculă sau grup de molecule similare, este numită situs de legare proteină-ligand sau situs activ. Liganzii sunt molecule care interacționează cu proteinele.
Specificitate ridicată Interacțiunea proteinei cu ligand este asigurată de complementaritatea structurii centrului activ cu structura ligandului.
Complementaritatea- aceasta este corespondența spațială și chimică a suprafețelor care interacționează. Centrul activ nu trebuie să corespundă doar spațial ligandului inclus în el, dar și legături (interacțiuni ionice, hidrogen și hidrofobe) trebuie să se formeze între grupările funcționale ale radicalilor incluși în centrul activ și ligand, care dețin ligandul. în centrul activ (Fig. 1.13 ).
Orez. 1.13. Interacțiunea complementară a proteinei cu ligand
Unii liganzi, atunci când sunt atașați la centrul activ al unei proteine, joacă un rol auxiliar în funcționarea proteinelor. Astfel de liganzi sunt numiți cofactori, iar proteinele care conțin o parte neproteică sunt numite proteine complexe(spre deosebire de proteinele simple, constând numai din partea proteică). Partea non-proteică, ferm conectată la proteină, se numește grupare prostetică. De exemplu, mioglobina, hemoglobina și citocromii conțin un grup protetic, hem, care conține un ion de fier, ferm atașat de centrul activ. Proteinele complexe care conțin hem se numesc hemoproteine.
Atunci când liganzii specifici sunt atașați de proteine, se manifestă funcția acestor proteine. Astfel, albumina, cea mai importantă proteină din plasma sanguină, își manifestă funcția de transport prin atașarea liganzilor hidrofobi, precum acizii grași, bilirubina, unele medicamente etc., la centrul activ (Fig. 1.14)
Liganzii care interacționează cu structura tridimensională a lanțului peptidic pot fi nu numai molecule organice și anorganice cu o moleculă scăzută, ci și macromolecule:
ADN (exemple cu proteine de legare la ADN discutate mai sus);
Polizaharide;
Orez. 1.14. Relația dintre genotip și fenotip
Structura primară unică a proteinelor umane, codificată în molecula de ADN, este realizată în celule sub forma unei conformații unice, a structurii centrului activ și a funcțiilor proteice.
În aceste cazuri, proteina recunoaște o regiune specifică a ligandului care este proporțională și complementară cu situsul de legare. Astfel, pe suprafața hepatocitelor există proteine receptor pentru hormonul insulină, care are și o structură proteică. Interacțiunea insulinei cu receptorul determină o modificare a conformației sale și activarea sistemelor de semnalizare, ducând la stocarea nutrienților în hepatocite după mese.
Prin urmare, Funcționarea proteinelor se bazează pe interacțiunea specifică a centrului activ al proteinei cu ligand.
2. Structura domeniului și rolul său în funcționarea proteinelor. Lanțurile polipeptidice lungi de proteine globulare se pliază adesea în mai multe regiuni compacte, relativ independente. Au o structură terțiară independentă, care amintește de cea a proteinelor globulare și sunt numite domenii. Datorită structurii de domeniu a proteinelor, structura lor terțiară este mai ușor de format.
În proteinele de domeniu, situsurile de legare a ligandului sunt adesea localizate între domenii. Astfel, tripsina este o enzimă proteolitică care este produsă de partea exocrină a pancreasului și este necesară pentru digestia proteinelor alimentare. Are o structură cu două domenii, iar centrul de legare a tripsinei cu ligandul său - proteina alimentară - este situat în șanțul dintre cele două domenii. În centrul activ, sunt create condițiile necesare pentru legarea eficientă a unui anumit situs al proteinei alimentare și hidroliza legăturilor sale peptidice.
Diferitele domenii dintr-o proteină se pot deplasa unul față de celălalt atunci când centrul activ interacționează cu ligand (Fig. 1.15).
Hexokinaza- o enzima care catalizeaza fosforilarea glucozei folosind ATP. Locul activ al enzimei este situat în despicatură dintre cele două domenii. Când hexokinaza se leagă de glucoză, domeniile din jurul acesteia se închid și substratul devine prins, unde are loc fosforilarea (vezi Fig. 1.15).
Orez. 1.15. Legarea domeniilor hexokinazei la glucoză
În unele proteine, domeniile îndeplinesc funcții independente prin legarea la diferiți liganzi. Astfel de proteine sunt numite multifuncționale.
3. Medicamentele sunt liganzi care afectează funcția proteinelor. Interacțiunea proteinelor cu liganzii este specifică. Cu toate acestea, datorită labilității conformaționale a proteinei și a centrului său activ, este posibil să se selecteze o altă substanță care ar putea interacționa și cu proteina din centrul activ sau din altă parte a moleculei.
Se numește o substanță similară ca structură cu un ligand natural analog structural al ligandului sau un ligand nenatural. De asemenea, interacționează cu proteina de la locul activ. Un analog structural al unui ligand poate îmbunătăți funcția proteinei (agonist),și reduceți-l (antagonist). Ligandul și analogii săi structurali concurează unul cu celălalt pentru legarea de proteină în același loc. Astfel de substanțe sunt numite modulatori competitivi(regulatori) ai funcţiilor proteinelor. Multe medicamente acționează ca inhibitori de proteine. Unele dintre ele sunt obținute prin modificarea chimică a liganzilor naturali. Inhibitorii funcțiilor proteinelor pot fi medicamente și otrăvuri.
Atropina este un inhibitor competitiv al receptorilor M-colinergici. Acetilcolina este un neurotransmitator pentru transmiterea impulsurilor nervoase prin sinapsele colinergice. Pentru a efectua excitația, acetilcolina eliberată în fanta sinaptică trebuie să interacționeze cu proteina receptoră a membranei postsinaptice. Două tipuri găsite receptori colinergici:
receptor M pe langa acetilcolina, interactioneaza selectiv cu muscarina (toxina agaric musca). M - receptorii colinergici sunt prezenți pe mușchii netezi și, atunci când interacționează cu acetilcolina, provoacă contracția acestora;
receptor H se leagă în mod specific de nicotină. Receptorii N-colinergici se găsesc la sinapsele mușchilor scheletici striați.
Inhibitor specific Receptorii M-colinergici este atropina. Se găsește în plantele belladonă și henbane.
Atropina are grupe funcționale similare ca structură cu acetilcolina și aranjarea lor spațială, prin urmare este un inhibitor competitiv al receptorilor M-colinergici. Având în vedere că legarea acetilcolinei de receptorii M-colinergici determină contracția mușchilor netezi, atropina este utilizată ca medicament care ameliorează spasmul acestora. (antispasmodic). Astfel, este cunoscută utilizarea atropinei pentru a relaxa mușchii ochilor la vizualizarea fundului de ochi, precum și pentru a ameliora spasmele în timpul colicilor gastrointestinale. Receptorii M-colinergici sunt prezenți și în sistemul nervos central (SNC), astfel încât dozele mari de atropină pot provoca o reacție nedorită a sistemului nervos central: agitație motorie și psihică, halucinații, convulsii.
Ditilina este un agonist competitiv al receptorilor H-colinergici, inhibă funcția sinapselor neuromusculare.
Sinapsele neuromusculare ale mușchilor scheletici conțin receptori H-colinergici. Interacțiunea lor cu acetilcolina duce la contracții musculare. În timpul unor operații chirurgicale, precum și în studiile endoscopice, sunt utilizate medicamente care provoacă relaxarea mușchilor scheletici. (relaxante musculare). Acestea includ ditilina, care este un analog structural al acetilcolinei. Se ataseaza de receptorii H-colinergici, dar spre deosebire de acetilcolina, este distrusa foarte lent de enzima acetilcolinesteraza. Ca urmare a deschiderii prelungite a canalelor ionice și a depolarizării persistente a membranei, conducerea impulsurilor nervoase este perturbată și are loc relaxarea musculară. Inițial, aceste proprietăți au fost descoperite în otravă de curare, motiv pentru care se numesc astfel de medicamente ca curare.
TEMA 1.3. DENATURAREA PROTEINELOR ŞI POSIBILITATEA RENATĂRII LOR SPONTANE
1. Deoarece conformația nativă a proteinelor este menținută din cauza interacțiunilor slabe, modificări ale compoziției și proprietăților mediului din jurul proteinei, expunerea la reactivi chimici și factori fizici provoacă o modificare a conformației acestora (proprietatea labilitatii conformaționale). Ruperea unui număr mare de legături duce la distrugerea conformației native și la denaturarea proteinelor.
Denaturarea proteinelor- aceasta este distrugerea conformatiei lor native sub influenta agentilor denaturanti, cauzata de ruperea legaturilor slabe care stabilizeaza structura spatiala a proteinei. Denaturarea este însoțită de distrugerea structurii unice tridimensionale și a centrului activ al proteinei și de pierderea activității sale biologice (Fig. 1.16).
Toate moleculele denaturate ale unei proteine dobândesc o conformație aleatorie care diferă de alte molecule ale aceleiași proteine. Radicalii de aminoacizi care formează centrul activ se dovedesc a fi distanțați spațial unul de celălalt, adică. locul de legare specific al proteinei cu ligand este distrus. În timpul denaturarii, structura primară a proteinelor rămâne neschimbată.
Aplicarea agenților de denaturare în cercetarea biologică și medicină.În studiile biochimice, înainte de a determina compușii cu greutate moleculară mică din materialul biologic, proteinele sunt de obicei îndepărtate mai întâi din soluție. Acidul tricloroacetic (TCA) este cel mai des folosit în acest scop. După adăugarea TCA la soluție, proteinele denaturate precipită și sunt ușor îndepărtate prin filtrare (Tabelul 1.1.)
În medicină, agenții de denaturare sunt adesea utilizați pentru sterilizarea instrumentelor și materialelor medicale în autoclave (agentul de denaturare este la temperatură ridicată) și ca antiseptice (alcool, fenol, cloramină) pentru tratarea suprafețelor contaminate care conțin microfloră patogenă.
2. Reactivarea proteinelor spontane- dovada determinismului structurii, conformaţiei şi funcţiei primare a proteinelor. Proteinele individuale sunt produse ale unei gene care au o secvență identică de aminoacizi și dobândesc aceeași conformație în celulă. Concluzia fundamentală că structura primară a unei proteine conține deja informații despre conformația și funcția sa a fost făcută pe baza capacității unor proteine (în special, ribonucleaza și mioglobina) de a se renativa spontan - restabili conformația lor nativă după denaturare.
Formarea structurilor proteice spațiale se realizează prin metoda auto-asamblarii - un proces spontan în care un lanț polipeptidic, care are o structură primară unică, tinde să adopte o conformație cu cea mai scăzută energie liberă în soluție. Capacitatea de a renativa proteinele care își păstrează structura primară după denaturare a fost descrisă într-un experiment cu enzima ribonuclează.
Ribonucleaza este o enzimă care descompune legăturile dintre nucleotidele individuale dintr-o moleculă de ARN. Această proteină globulară are un lanț polipeptidic, a cărui structură terțiară este stabilizată de multe legături slabe și patru legături disulfurice.
Tratamentul ribonucleazei cu uree, care rupe legăturile de hidrogen din moleculă și un agent reducător, care rupe legăturile disulfurice, duce la denaturarea enzimei și la pierderea activității acesteia.
Îndepărtarea agenților denaturanți prin dializă duce la restabilirea conformației și funcției proteinelor, de exemplu. a renaște. (Fig. 1.17).
Orez. 1.17. Denaturarea și renativarea ribonucleazei
A - conformația nativă a ribonucleazei, în structura terțiară a cărei patru legături disulfurice; B - moleculă de ribonuclează denaturată;
B - moleculă de ribonuclează reactivată cu structură și funcție restaurate
1. Completați tabelul 1.2.
Tabelul 1.2. Clasificarea aminoacizilor în funcție de polaritatea radicalilor
2. Scrieți formula tetrapeptidei:
Asp - Pro - Fen - Liz
a) evidențiați grupările repetate din peptidă care formează coloana vertebrală peptidică și grupările variabile reprezentate de radicalii aminoacizi;
b) etichetați N- și C-terminali;
c) evidenţiază legăturile peptidice;
d) scrieți o altă peptidă formată din aceiași aminoacizi;
e) numărați numărul de variante posibile ale unei tetrapeptide cu o compoziție similară de aminoacizi.
3. Explicați rolul structurii primare a proteinelor folosind exemplul unei analize comparative a doi hormoni peptidici similari din punct de vedere structural și apropiați din punct de vedere evolutiv ai neurohipofizei mamiferelor - oxitocina și vasopresina (Tabelul 1.3).
Tabelul 1.3. Structura și funcțiile oxitocinei și vasopresinei
Pentru aceasta:
a) comparați compoziția și secvența de aminoacizi a două peptide;
b) găsiți asemănarea structurii primare a celor două peptide și asemănarea acțiunii lor biologice;
c) găsiți diferențe în structura a două peptide și diferențe în funcțiile acestora;
d) trageți o concluzie despre influența structurii primare a peptidelor asupra funcțiilor acestora.
4. Descrieți principalele etape de formare a conformației proteinelor globulare (structuri secundare, terțiare, conceptul de structură supersecundară). Indicați tipurile de legături implicate în formarea structurilor proteice. Ce radicali de aminoacizi pot participa la formarea interacțiunilor hidrofobe, ionice, legături de hidrogen.
Dă exemple.
5. Definiți conceptul de „labilitatea conformațională a proteinelor”, indicați motivele existenței și semnificația acestuia.
6. Extindeți semnificația următoarei fraze: „Funcționarea proteinelor se bazează pe interacțiunea lor specifică cu ligandul”, folosind termenii și explicând sensul lor: conformație proteinei, centru activ, ligand, complementaritate, funcție proteică.
7. Folosind un exemplu, explicați ce domenii sunt și care este rolul lor în funcționarea proteinelor.
SARCINI DE AUTOCONTROL
1. Meci.
Grupa funcțională în radicalul aminoacid:
A. Gruparea carboxil B. Gruparea hidroxil C Gruparea guanidină D. Gruparea tiol E. Gruparea amino
2. Alege raspunsurile corecte.
Aminoacizii cu radicali polari neîncărcați sunt:
A. Cis B. Asn
B. Glu G. Trei
3. Alege raspunsurile corecte.
Radicali de aminoacizi:
A. Asigură specificitatea structurii primare B. Participa la formarea structurii terțiare
B. Situate pe suprafața proteinei, ele influențează solubilitatea acesteia D. Formează centrul activ
D. Participa la formarea legăturilor peptidice
4. Alege raspunsurile corecte.
Interacțiunile hidrofobe se pot forma între radicalii de aminoacizi:
A. Tre Lay B. Pro Trei
B. Met Ile G. Tir Ala D. Val Fen
5. Alege raspunsurile corecte.
Între radicalii de aminoacizi se pot forma legături ionice:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn apr
6. Alege raspunsurile corecte.
Legăturile de hidrogen se pot forma între radicalii de aminoacizi:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Meci.
Tipul de legătură implicat în formarea structurii proteinei:
A. Structura primară B. Structura secundară
B. Structura terţiară
D. Structura suprasecundară E. Conformaţia.
1. Legături de hidrogen între atomii scheletului peptidic
2. Legături slabe între grupările funcționale ale radicalilor aminoacizi
3. Legături între grupările α-amino și α-carboxil ale aminoacizilor
8. Alege raspunsurile corecte. Tripsina:
A. Enzima proteolitică B. Conține două domenii
B. Hidrolizează amidonul
D. Site-ul activ este situat între domenii. D. Constă din două lanțuri polipeptidice.
9. Alege raspunsurile corecte. Atropina:
A. Neurotransmițător
B. Analog structural al acetilcolinei
B. Interacționează cu receptorii H-colinergici
D. Intareste conducerea impulsurilor nervoase prin sinapsele colinergice
D. Inhibitor competitiv al receptorilor M-colinergici
10. Alegeți afirmațiile corecte. În proteine:
A. Structura primară conține informații despre structura site-ului său activ
B. Centrul activ se formează la nivelul structurii primare
B. Conformația este fixată rigid prin legături covalente
D. Situl activ poate interacționa cu un grup de liganzi similari
datorită labilităţii conformaţionale a proteinelor D. Modificările mediului pot afecta afinitatea activului
centru la ligand
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
TERMENI ȘI CONCEPTE DE BAZĂ
1. Proteine, polipeptide, aminoacizi
2. Structuri proteice primare, secundare, terțiare
3. Conformația, conformația proteinei native
4. Legături covalente și slabe în proteine
5. Labilitatea conformațională
6. Sit activ proteic
7. Liganzi
8. Plierea proteinelor
9. Analogi structurali ai liganzilor
10. Proteine de domeniu
11. Proteine simple și complexe
12. Denaturarea proteinelor, agenți de denaturare
13. Reactivarea proteinelor
Rezolva probleme
„Organizarea structurală a proteinelor și baza funcționării lor”
1. Funcția principală a proteinei - hemoglobina A (HbA) este transportul oxigenului către țesuturi. În populația umană, sunt cunoscute forme multiple ale acestei proteine cu proprietăți și funcții modificate - așa-numitele hemoglobine anormale. De exemplu, hemoglobina S, găsită în globulele roșii ale pacienților cu siclemie (HbS), s-a dovedit a avea o solubilitate scăzută în condiții de presiune parțială scăzută a oxigenului (cum este cazul sângelui venos). Acest lucru duce la formarea de agregate ale acestei proteine. Proteina își pierde funcția, precipită, iar celulele roșii din sânge devin neregulate (unele dintre ele formează o formă de seceră) și sunt distruse mai repede decât de obicei în splină. Ca urmare, se dezvoltă anemia cu celule falciforme.
Singura diferență în structura primară a HbA a fost găsită în regiunea N-terminală a lanțului β al hemoglobinei. Comparați regiunile N-terminale ale catenei β și arată cum modificările în structura primară a proteinei îi afectează proprietățile și funcțiile.
Pentru aceasta:
a) scrieți formulele aminoacizilor prin care HbA diferă și comparați proprietățile acestor aminoacizi (polaritate, sarcină).
b) trageți o concluzie despre motivul scăderii solubilității și perturbării transportului oxigenului în țesuturi.
2. Figura prezintă o diagramă a structurii unei proteine care are un centru de legare cu un ligand (centru activ). Explicați de ce proteina este selectivă în alegerea ligandului. Pentru aceasta:
a) amintiți-vă care este centrul activ al unei proteine și luați în considerare structura centrului activ al proteinei prezentată în figură;
b) scrieți formulele radicalilor aminoacizi care alcătuiesc centrul activ;
c) trageți un ligand care ar putea interacționa în mod specific cu situsul activ al proteinei. Indicați pe el grupele funcționale care pot forma legături cu radicalii aminoacizi care alcătuiesc centrul activ;
d) indicați tipurile de legături care apar între radicalii ligand și aminoacizi ai situsului activ;
e) explicați pe ce se bazează specificitatea interacțiunii proteină-ligand.
3.
Figura prezintă locul activ al proteinei și al mai multor liganzi.
Determinați care ligand este cel mai probabil să interacționeze cu situsul activ al proteinei și de ce.
Ce tipuri de legături apar în timpul formării unui complex proteină-ligand?
4. Analogii structurali ai liganzilor proteici naturali pot fi utilizați ca medicamente pentru a modifica activitatea proteinelor.
Acetilcolina este un mediator al transmiterii excitației la sinapsele neuromusculare. Când acetilcolina interacționează cu proteinele - receptorii membranei postsinaptice a mușchilor scheletici, canalele ionice se deschid și are loc contracția musculară. Ditilin este un medicament folosit în unele operații pentru relaxarea mușchilor, deoarece întrerupe transmiterea impulsurilor nervoase prin sinapsele neuromusculare. Explicați mecanismul de acțiune al ditilinei ca relaxant muscular. Pentru aceasta:
a) scrieți formulele acetilcolinei și ditilinei și comparați structurile acestora;
b) descrieți mecanismul efectului relaxant al ditilinei.
5. În unele boli, temperatura corpului pacientului crește, ceea ce este considerat o reacție de protecție a corpului. Cu toate acestea, temperaturile ridicate sunt dăunătoare pentru proteinele corpului. Explicați de ce la temperaturi peste 40 °C funcția proteinelor este perturbată și apare o amenințare la adresa vieții umane. Pentru a face acest lucru, amintiți-vă:
1) Structura proteinelor și legăturile care îi mențin structura în conformația nativă;
2) Cum se modifică structura și funcția proteinelor odată cu creșterea temperaturii?;
3) Ce este homeostazia și de ce este importantă pentru menținerea sănătății umane.
Unitate modulară 2 PROTEINE OLIGOMERICE CA ȚINTE ALE INFLUENȚELOR REGLATORII. DIVERSITATEA STRUCTURALA SI FUNCTIONALA A PROTEINELOR. METODE DE SEPARARE ȘI PURIFICARE A PROTEINELOR
Obiective de învățare A fi capabil să:
1. Utilizați cunoștințele despre caracteristicile structurii și funcțiilor proteinelor oligomerice pentru a înțelege mecanismele adaptative de reglare a funcțiilor acestora.
2. Explicați rolul chaperonelor în sinteza și menținerea conformației proteinelor în condiții celulare.
3. Explicați varietatea manifestărilor vieții prin varietatea structurilor și funcțiilor proteinelor sintetizate în organism.
4. Analizați relația dintre structura proteinelor și funcția lor folosind exemple de comparare a hemoproteinelor înrudite - mioglobină și hemoglobină, precum și reprezentanți ai celor cinci clase de proteine din familia imunoglobulinelor.
5. Aplicarea cunoștințelor despre particularitățile proprietăților fizice și chimice ale proteinelor pentru a selecta metode de purificare a acestora din alte proteine și impurități.
6. Interpretați rezultatele compoziției cantitative și calitative a proteinelor plasmatice pentru a confirma sau clarifica diagnosticul clinic.
Știi:
1. Caracteristici ale structurii proteinelor oligomerice și mecanisme adaptative de reglare a funcțiilor acestora folosind exemplul hemoglobinei.
2. Structura și funcțiile chaperonelor și importanța acestora pentru menținerea conformației native a proteinelor în condiții celulare.
3. Principii de combinare a proteinelor în familii pe baza asemănării conformației și funcțiilor lor folosind exemplul imunoglobulinelor.
4. Metode de separare a proteinelor pe baza caracteristicilor proprietăților lor fizico-chimice.
5. Electroforeza plasmei sanguine ca metodă de evaluare a compoziției calitative și cantitative a proteinelor.
TEMA 1.4. CARACTERISTICI ALE STRUCTURII ȘI FUNCȚIONĂRII PROTEINELOR OLIGOMERICE CU EXEMPLU DE HEMOGLOBINĂ
1. Multe proteine conțin mai multe lanțuri polipeptidice. Astfel de proteine sunt numite oligomerice,și lanțuri individuale - protomeri. Protomerii din proteinele oligomerice sunt legați prin multe legături slabe necovalente (hidrofobe, ionice, hidrogen). Interacţiune
protomerii se realizează datorită complementaritatea suprafețele lor de contact.
Numărul de protomeri din proteinele oligomerice poate varia foarte mult: hemoglobina conține 4 protomeri, enzima aspartat aminotransferaza are 12 protomeri, iar proteina virusului mozaicului tutunului conține 2120 protomeri legați prin legături necovalente. În consecință, proteinele oligomerice pot avea greutăți moleculare foarte mari.
Interacțiunea unui protomer cu alții poate fi considerată un caz special de interacțiune proteină-ligand, deoarece fiecare protomer servește ca ligand pentru alți protomeri. Numărul și metoda de unire a protomerilor într-o proteină se numesc structura proteinelor cuaternare.
Proteinele pot conține protomeri cu structuri identice sau diferite, de exemplu, homodimerii sunt proteine care conțin doi protomeri identici, iar heterodimerii sunt proteine care conțin doi protomeri diferiți.
Dacă proteinele conțin protomeri diferiți, atunci se pot forma pe ele centre de legare cu liganzi diferiți, care diferă ca structură. Când un ligand se leagă de situsul activ, se manifestă funcția acestei proteine. Un centru situat pe un protomer diferit se numește alosteric (diferit de cel activ). Contactarea ligand sau efector alosteric,îndeplineşte o funcţie de reglementare (Fig. 1.18). Interacțiunea centrului alosteric cu efectorul provoacă modificări conformaționale în structura întregii proteine oligomerice datorită labilității conformaționale a acesteia. Acest lucru afectează afinitatea situsului activ pentru un ligand specific și reglează funcția acelei proteine. O schimbare în conformația și funcția tuturor protomerilor în timpul interacțiunii unei proteine oligomere cu cel puțin un ligand se numește modificări conformaționale cooperative. Se numesc efectori care îmbunătățesc funcția proteinelor activatori,și efectori care îi inhibă funcția - inhibitori.
Astfel, proteinele oligomerice, precum și proteinele cu structură de domeniu, au o nouă proprietate în comparație cu proteinele monomerice - capacitatea de a regla alosteric funcțiile (reglare prin atașarea diferiților liganzi la proteină). Acest lucru poate fi văzut comparând structurile și funcțiile a două proteine complexe strâns legate, mioglobina și hemoglobina.
Orez. 1.18. Schema structurii unei proteine dimerice
2. Formarea structurilor spațiale și funcționarea mioglobinei.
Mioglobina (Mb) este o proteină care se găsește în mușchii roșii, a cărei funcție principală este de a crea rezerve de O 2 necesare lucrului muscular intens. Mb este o proteină complexă care conține o parte proteică - apoMb și o parte neproteică - hem. Structura primară a apoMB determină conformația sa globulară compactă și structura centrului activ, de care este atașată partea neproteică a mioglobinei, hem. Oxigenul care vine din sânge către mușchi se leagă de hemii Fe+2 din mioglobină. Mb este o proteină monomerică care are o afinitate foarte mare pentru O 2, prin urmare eliberarea de oxigen de către mioglobină are loc numai în timpul muncii musculare intense, când presiunea parțială a O 2 scade brusc.
Formarea conformaţiei Mv.În mușchii roșii, pe ribozomi în timpul translației, se sintetizează structura primară a MB, reprezentată de o secvență specifică de 153 de resturi de aminoacizi. Structura secundară a lui Mb conține opt elice α, numite cu litere latine de la A la H, între care există regiuni nehelicoidale. Structura terțiară a lui Mb are forma unui globul compact, în adâncitura căruia se află centrul activ între elicele α F și E (Fig. 1.19).
Orez. 1.19. Structura mioglobinei
3. Caracteristici ale structurii și funcționării centrului activ al MV. Centrul activ al Mb este format predominant din radicali de aminoacizi hidrofobi, distanțați larg unul de celălalt în structura primară (de exemplu, Tri 3 9 și Fen 138) Puțin solubili în apă liganzii - hem și O 2 - se atașează la centrul activ. Hemul este un ligand specific al apoMB (Fig. 1.20), a cărui bază este alcătuită din patru inele pirol conectate prin punți metanilice; în centru se află un atom de Fe+ 2 legat de atomii de azot ai inelelor pirol prin patru legături de coordonare. În centrul activ al Mb, pe lângă radicalii de aminoacizi hidrofobi, există și reziduuri a doi aminoacizi cu radicali hidrofili - Gis E 7(Gis 64) și GIS F 8(His 93) (Fig. 1.21).
Orez. 1.20. Structura hemului - partea neproteică a mioglobinei și hemoglobinei
Orez. 1.21. Localizarea hemului și a O2 în locul activ al apomioglobinei și al protomerilor hemoglobinei
Hemul este legat covalent de His F8 printr-un atom de fier. O 2 se atașează de fierul de călcat pe cealaltă parte a planului hem. Lui E 7 este necesar pentru orientarea corectă a O 2 și facilitează adăugarea de oxigen la Fe + 2 hem
GIS F 8 formează o legătură de coordonare cu Fe+ 2 și fixează ferm hemul în centrul activ. Gis E 7 necesar pentru orientarea corectă în centrul activ al altui ligand - O 2 în timpul interacțiunii acestuia cu Fe + 2 hem. Micromediul hemului creează condiții pentru legarea puternică, dar reversibilă, a O2 de Fe + 2 și împiedică intrarea apei în locul activ hidrofob, ceea ce poate duce la oxidarea acestuia la Fe + 3.
Structura monomerică a Mb și centrul său activ determină afinitatea ridicată a proteinei pentru O2.
4. Structura oligomerică a Hb și reglarea afinității Hb față de liganzii O2. Hemoglobinele umane- o familie de proteine, precum mioglobina, legate de proteine complexe (hemoproteine). Au o structură tetramerică și conțin două lanțuri α, dar diferă prin structura celorlalte două lanțuri polipeptidice (lanțuri 2α, 2x). Structura celui de-al doilea lanț polipeptidic determină caracteristicile funcționării acestor forme de Hb. Aproximativ 98% din hemoglobina din globulele roșii ale unui adult este hemoglobina A(2α-, 2p-lanțuri).
În timpul dezvoltării fetale, funcționează două tipuri principale de hemoglobine: Hb embrionară(2α, 2ε), care se găsește în stadiile incipiente ale dezvoltării fetale și hemoglobina F (fetală)- (2α, 2γ), care înlocuiește hemoglobina fetală precoce în luna a șasea de dezvoltare intrauterină și numai după naștere este înlocuită cu Hb A.
HB A este o proteină legată de mioglobina (MB) găsită în globulele roșii umane adulte. Structura protomerilor săi individuali este similară cu cea a mioglobinei. Structurile secundare și terțiare ale mioglobinei și protomerilor hemoglobinei sunt foarte asemănătoare, în ciuda faptului că în structura primară a lanțurilor lor polipeptidice doar 24 de resturi de aminoacizi sunt identice (structura secundară a protomerilor hemoglobinei, ca și mioglobina, conține opt α-helice, desemnată prin litere latine de la A la H , iar structura terţiară are forma unui globule compact). Dar, spre deosebire de mioglobina, hemoglobina are o structură oligomerică, constând din patru lanțuri polipeptidice conectate prin legături necovalente (Figura 1.22).
Fiecare protomer Hb este asociat cu o parte non-proteică - hem și protomeri vecini. Legătura părții proteice a Hb cu hem este similară cu cea a mioglobinei: în centrul activ al proteinei, părțile hidrofobe ale hemului sunt înconjurate de radicali hidrofobi de aminoacizi, cu excepția His F 8 și His E 7, care sunt situate pe ambele părți ale planului hem și joacă un rol similar în funcționarea proteinei și în legarea acesteia cu oxigenul (vezi structura mioglobinei).
Orez. 1.22. Structura oligomerică a hemoglobinei
In afara de asta, Gis E 7 realizează un important rol suplimentarîn funcționarea Nv. Hemul liber are o afinitate de 25.000 de ori mai mare pentru CO decât pentru O2. CO se formează în cantități mici în organism și, având în vedere afinitatea sa mare pentru hem, ar putea perturba transportul de O 2 necesar vieții celulare. Totuși, în compoziția hemoglobinei, afinitatea hemului pentru monoxidul de carbon depășește de numai 200 de ori afinitatea pentru O 2 datorită prezenței His E 7 în centrul activ. Restul acestui aminoacid creează condiții optime pentru legarea hemului de O2 și slăbește interacțiunea hemului cu CO.
5. Funcția principală a HB este transportul O2 de la plămâni la țesuturi. Spre deosebire de mioglobina monomerică, care are o afinitate foarte mare pentru O2 și îndeplinește funcția de stocare a oxigenului în mușchii roșii, structura oligomerică a hemoglobinei oferă:
1) saturarea rapidă a HB cu oxigen în plămâni;
2) capacitatea HB de a elibera oxigen în ţesuturi la o presiune parţială relativ mare de O2 (20-40 mm Hg);
3) posibilitatea de reglare a afinității Hb față de O 2.
6. Modificările cooperante în conformația protomerilor hemoglobinei accelerează legarea O 2 în plămâni și eliberarea acestuia în țesuturi. În plămâni, presiunea parțială ridicată a O2 promovează legarea acestuia de Hb în situsul activ a patru protomeri (2α și 2β). Centrul activ al fiecărui protomer, ca și în mioglobină, este situat între două elice α (F și E) într-un buzunar hidrofob. Conține o parte non-proteică - hem, atașată de partea proteică prin multe interacțiuni hidrofobe slabe și o legătură puternică între Fe 2 + hem și His F 8 (vezi Fig. 1.21).
În deoxihemoglobină, datorită acestei legături cu His F 8, atomul de Fe 2 + iese din planul hem către histidină. Legarea O2 la Fe2+ are loc pe cealaltă parte a hemului în regiunea His E7 folosind o singură legătură de coordonare liberă. His E 7 oferă condiții optime pentru legarea O 2 de fierul hem.
Adăugarea de O 2 la atomul de Fe + 2 al unui protomer determină mișcarea acestuia în planul hem, urmată de reziduul de histidină asociat cu acesta
Orez. 1.23. Modificarea conformației protomerului hemoglobinei atunci când este combinat cu O2
Acest lucru duce la o modificare a conformației tuturor lanțurilor polipeptidice datorită labilității lor conformaționale. Modificarea conformației altor lanțuri facilitează interacțiunea acestora cu moleculele de O 2 ulterioare.
A patra moleculă de O 2 se atașează de hemoglobină de 300 de ori mai ușor decât prima (Fig. 1.24).
Orez. 1.24. Modificări cooperante în conformația protomerilor hemoglobinei în timpul interacțiunii sale cu O2
În țesuturi, fiecare moleculă de O2 ulterioară este scindată mai ușor decât cea anterioară, tot datorită modificărilor cooperante în conformația protomerilor.
7. CO 2 şi H+, formate în timpul catabolismului substanţelor organice, reduc afinitatea hemoglobinei pentru O 2 proporţional cu concentraţia acestora. Energia necesară pentru funcționarea celulelor este produsă în primul rând în mitocondrii în timpul oxidării substanțelor organice folosind O 2 eliberat din plămâni de hemoglobină. Ca urmare a oxidării substanţelor organice se formează produşii finali ai descompunerii acestora: CO 2 şi K 2 O, a căror cantitate este proporţională cu intensitatea proceselor de oxidare în curs.
CO 2 difuzează din celule în sânge și pătrunde în celulele roșii din sânge, unde, sub acțiunea enzimei carbanhidraze, este transformat în acid carbonic. Acest acid slab se disociază într-un proton și un ion de bicarbonat.
H+ sunt capabili să se alăture radicalilor Săi 14 6 în lanțurile α și β ale hemoglobinei, adică în zone îndepărtate de hem. Protonarea hemoglobinei reduce afinitatea acesteia pentru O 2, favorizează eliminarea O 2 din oxiHb, formarea deoxiHb și crește aportul de oxigen către țesuturi proporțional cu numărul de protoni formați (Fig. 1.25).
O creștere a cantității de oxigen eliberat în funcție de creșterea concentrației de H+ în celulele roșii din sânge se numește efect Bohr (numit după fiziologul danez Christian Bohr, care a descoperit primul acest efect).
În plămâni, o presiune parțială mare a oxigenului promovează legarea acestuia la deoxiHb, ceea ce reduce afinitatea proteinei pentru H +. Protonii eliberați sub acțiunea acidului carbonic reacționează cu bicarbonații pentru a forma CO 2 și H 2 O
Orez. 1.25. Dependența afinității Hb pentru O 2 de concentrația de CO 2 și protoni (efectul Bohr):
A- influenţa concentraţiei de CO 2 şi H+ asupra eliberării de O 2 din complexul cu HB (efect Bohr); B- oxigenarea deoxihemoglobinei în plămâni, formarea și eliberarea de CO2.
CO2 rezultat intră în spațiul alveolar și este îndepărtat cu aerul expirat. Astfel, cantitatea de oxigen eliberată de hemoglobină în țesuturi este reglată de produșii catabolismului substanțelor organice: cu cât descompunerea substanțelor este mai intensă, de exemplu în timpul exercițiului fizic, cu atât concentrația de CO 2 și H + este mai mare și cu atât mai mult oxigen. ţesuturile primesc ca urmare a scăderii afinităţii Hb pentru O 2.
8. Reglarea alosterică a afinității Hb pentru O2 de către ligand - 2,3-bisfosfoglicerat.În eritrocite, ligandul alosteric al hemoglobinei, 2,3-bisfosfogliceratul (2,3-BPG), este sintetizat din produsul oxidării glucozei - 1,3-bisfosfoglicerat. În condiții normale, concentrația de 2,3-BPG este ridicată și comparabilă cu concentrația de Hb. 2,3-BPG are o sarcină negativă puternică de -5.
Bisfosfogliceratul din capilarele tisulare, care se leagă de deoxihemoglobină, crește eliberarea de oxigen în țesuturi, reducând afinitatea Hb pentru O2.
În centrul moleculei de hemoglobină tetramerică se află o cavitate. Este format din resturile de aminoacizi ale tuturor celor patru protomeri (vezi Fig. 1.22). În capilarele tisulare, protonarea Hb (efectul Bohr) duce la ruperea legăturii dintre fierul hem și O2. Într-o moleculă
deoxihemoglobina, comparativ cu oxihemoglobina, apar legături ionice suplimentare care leagă protomerii, drept urmare dimensiunile cavității centrale cresc față de oxihemoglobina. Cavitatea centrală este locul de atașare a 2,3-BPG la hemoglobină. Datorită diferenței de dimensiune a cavității centrale, 2,3-BPG se poate atașa doar la deoxihemoglobină.
2,3-BPG interacționează cu hemoglobina într-un loc îndepărtat de centrii activi ai proteinei și aparține alosterică liganzi (reglatori), iar cavitatea centrală a Hb este centru alosteric. 2,3-BPG are o sarcină negativă puternică și interacționează cu cinci grupe încărcate pozitiv ale celor două lanțuri β ale Hb: gruparea α-amino N-terminală a lui Val și radicalii Lys 82 His 143 (Fig. 1.26).
Orez. 1.26. BPG în cavitatea centrală a deoxihemoglobinei
BPG se leagă de trei grupuri încărcate pozitiv pe fiecare catenă β.
În capilarele tisulare, deoxihemoglobina rezultată interacționează cu 2,3-BPG și se formează legături ionice între radicalii încărcați pozitiv ai lanțurilor β și ligandul încărcat negativ, care modifică conformația proteinei și reduc afinitatea Hb pentru O2. . O scădere a afinității Hb pentru O 2 contribuie la o eliberare mai eficientă a O 2 în țesut.
În plămâni, la presiune parțială mare, oxigenul interacționează cu Hb, unind fierul hem; în acest caz, conformația proteinei se modifică, cavitatea centrală scade și 2,3-BPG este deplasat din centrul alosteric.
Astfel, proteinele oligomerice au proprietăți noi în comparație cu proteinele monomerice. Atașarea liganzilor la situsuri
distanță spațial unul de celălalt (alosteric), poate provoca modificări conformaționale în întreaga moleculă proteică. Datorită interacțiunii cu liganzii de reglare, are loc o modificare a conformației și adaptarea funcției moleculei proteice la schimbările de mediu.
TEMA 1.5. MENȚINEREA CONFORMĂȚII NATIVE A PROTEINELOR ÎN CONDIȚII CELULARE
În celule, în timpul sintezei lanțurilor polipeptidice, transportul acestora prin membrane către părțile corespunzătoare ale celulei, în timpul procesului de pliere (formarea conformației native) și în timpul asamblării proteinelor oligomerice, precum și în timpul funcționării lor, intermediare. În structura proteinei apar conformații instabile, predispuse la agregare. Radicalii hidrofobi, de obicei ascunși în interiorul moleculei proteice în conformația nativă, apar la suprafață într-o conformație instabilă și tind să se combine cu grupuri de alte proteine care sunt slab solubile în apă. În celulele tuturor organismelor cunoscute, s-au găsit proteine speciale care asigură plierea optimă a proteinelor celulare, stabilizează conformația lor nativă în timpul funcționării și, cel mai important, mențin structura și funcțiile proteinelor intracelulare atunci când homeostazia este perturbată. Aceste proteine sunt numite "socitori" care înseamnă „dădacă” în franceză.
1. Chaperonele moleculare și rolul lor în prevenirea denaturarii proteinelor.
Chaperonele (CH) sunt clasificate în funcție de masa subunităților lor. Chaperonele cu greutate moleculară mare au o masă de la 60 la 110 kDa. Dintre acestea, trei clase au fost cele mai studiate: Sh-60, Sh-70 și Sh-90. Fiecare clasă include o familie de proteine înrudite. Astfel, Sh-70 include proteine cu o greutate moleculară de la 66 la 78 kDa. Chaperonele cu greutate moleculară mică au o greutate moleculară de la 40 la 15 kDa.
Printre însoțitori se numără constitutiv proteine, a căror sinteză bazală ridicată nu depinde de efectele stresului asupra celulelor organismului și inductibil, a cărui sinteză în condiții normale este slabă, dar crește brusc sub stres. Chaperonele inductibile sunt numite și „proteine de șoc termic”, deoarece au fost descoperite pentru prima dată în celulele expuse la temperaturi ridicate. În celule, datorită concentrației mari de proteine, reactivarea spontană a proteinelor parțial denaturate este dificilă. Sh-70 poate preveni apariția denaturarii și poate ajuta la restabilirea conformației native a proteinelor. Însoțitori moleculari-70- o clasă foarte conservată de proteine găsite în toate părțile celulei: citoplasmă, nucleu, reticul endoplasmatic, mitocondrii. La capătul carboxil al lanțului polipeptidic unic Ш-70 există o regiune care este un șanț capabil să interacționeze cu peptide de lungime
de la 7 la 9 resturi de aminoacizi îmbogățite cu radicali hidrofobi. Astfel de regiuni din proteinele globulare apar aproximativ la fiecare 16 aminoacizi. Sh-70 este capabil să protejeze proteinele de inactivarea temperaturii și să restabilească conformația și activitatea proteinelor parțial denaturate.
2. Rolul chaperonelor în plierea proteinelor.În timpul sintezei proteinelor pe ribozom, regiunea N-terminală a polipeptidei este sintetizată înaintea celei C-terminale. Pentru a forma conformația nativă, este necesară secvența completă de aminoacizi a proteinei. În procesul de sinteză a proteinelor, chaperonele-70, datorită structurii centrului lor activ, sunt capabile să închidă zonele polipeptidei predispuse la agregare, îmbogățite în radicali de aminoacizi hidrofobi până la finalizarea sintezei (Figura 1.27, A). ).
Orez. 1.27. Participarea chaperonelor la plierea proteinelor
A - participarea chaperonelor-70 la prevenirea interacțiunilor hidrofobe între secțiuni ale polipeptidei sintetizate; B - formarea conformației native a proteinei din complexul de chaperonă
Multe proteine cu molecule înalte care au o conformație complexă, cum ar fi o structură de domeniu, se pliază într-un spațiu special format din Sh-60. Ш-60 funcţionează ca un complex oligomeric format din 14 subunităţi. Ele formează două inele goale, fiecare dintre ele constând din șapte subunități, aceste inele sunt conectate între ele. Fiecare subunitate Sh-60 este formată din trei domenii: apical (apical), îmbogățit cu radicali hidrofobi orientați spre cavitatea inelului, intermediar și ecuatorial (Fig. 1.28).
Orez. 1.28. Structura complexului de chaperonine constând din 14 Ш-60
A - vedere laterală; B - vedere de sus
Proteinele sintetizate, care au elemente pe suprafața caracteristice moleculelor desfășurate, în special radicalii hidrofobi, intră în cavitatea inelelor chaperone. În mediul specific acestor cavități se caută conformații posibile până când se găsește singura care este cea mai favorabilă energetic (Fig. 1.27, B). Formarea conformațiilor și eliberarea proteinelor este însoțită de hidroliza ATP în regiunea ecuatorială. De obicei, o astfel de pliere dependentă de însoțitor necesită o cantitate semnificativă de energie.
Pe lângă participarea la formarea structurii tridimensionale a proteinelor și la renativarea proteinelor parțial denaturate, însoțitorii sunt, de asemenea, necesari pentru apariția unor procese fundamentale precum asamblarea proteinelor oligomerice, recunoașterea și transportul proteinelor denaturate în lizozomi, transportul proteinelor prin membrane și participarea la reglarea activității complexelor proteice.
TEMA 1.6. VARIETATE DE PROTEINE. FAMILIILE DE PROTEINE: EXEMPLU DE IMUNOGLOBULINE
1. Proteinele joacă un rol decisiv în viața celulelor individuale și a întregului organism multicelular, iar funcțiile lor sunt surprinzător de diverse. Acest lucru este determinat de caracteristicile structurii primare și conformațiilor proteinelor, de structura unică a centrului activ și de capacitatea de a lega liganzi specifici.
Doar o fracțiune foarte mică din toate variantele posibile de lanțuri peptidice poate adopta o structură spațială stabilă; majoritate
dintre ele pot lua multe conformații cu aproximativ aceeași energie Gibbs, dar cu proprietăți diferite. Structura primară a majorității proteinelor cunoscute selectate prin evoluția biologică asigură o stabilitate excepțională a uneia dintre conformații, ceea ce determină caracteristicile funcționării acestei proteine.
2. Familii de proteine.În cadrul aceleiași specii biologice, substituțiile de resturi de aminoacizi pot duce la apariția diferitelor proteine care îndeplinesc funcții înrudite și au secvențe de aminoacizi omoloage. Astfel de proteine înrudite au conformații surprinzător de similare: numărul și pozițiile relative ale elicelor a și/sau structurilor p și majoritatea spirelor și curburilor lanțurilor polipeptidice sunt similare sau identice. Proteinele cu regiuni omoloage ale lanțului polipeptidic, conformație similară și funcții înrudite sunt clasificate în familii de proteine. Exemple de familii de proteine: serin proteinaze, familia imunoglobulinelor, familia mioglobinelor.
Serin proteinaze- o familie de proteine care îndeplinesc funcția de enzime proteolitice. Acestea includ enzime digestive - chimotripsina, tripsina, elastaza si multi factori de coagulare a sangelui. Aceste proteine au aminoacizi identici în 40% din pozițiile lor și o conformație foarte asemănătoare (Fig. 1.29).
Orez. 1.29. Structuri spațiale ale elastazei (A) și chimotripsinei (B)
Unele substituții de aminoacizi au condus la modificări ale specificității substratului acestor proteine și la apariția diversității funcționale în cadrul familiei.
3. Familia imunoglobulinelor.În funcționarea sistemului imunitar, proteinele superfamiliei imunoglobulinelor joacă un rol imens, care include trei familii de proteine:
Anticorpi (imunoglobuline);
receptorii limfocitelor T;
Proteine ale complexului major de histocompatibilitate - MHC clasele 1 și 2 (Complex major de histocompatibilitate).
Toate aceste proteine au o structură de domeniu, constau din domenii asemănătoare imunologice omoloage și îndeplinesc funcții similare: interacționează cu structuri străine, fie dizolvate în sânge, limfă sau lichid intercelular (anticorpi), fie situate la suprafața celulelor (proprii sau străin).
4. Anticorpi- proteine specifice produse de limfocitele B ca răspuns la intrarea unei structuri străine în organism, numite antigen.
Caracteristicile structurii anticorpilor
Cele mai simple molecule de anticorpi constau din patru lanțuri polipeptidice: două ușoare identice - L, care conțin aproximativ 220 de aminoacizi, și două grele identice - H, constând din 440-700 de aminoacizi. Toate cele patru lanțuri din molecula de anticorp sunt conectate prin multe legături necovalente și patru legături disulfurice (Fig. 1.30).
Lanțurile ușor de anticorpi constau din două domenii: un domeniu variabil (VL), situat în regiunea N-terminală a lanțului polipeptidic și un domeniu constant (CL), situat la capătul C-terminal. Lanțurile grele au de obicei patru domenii: o variabilă (VH), situată la capătul N-terminal și trei domenii constante (CH1, CH2, CH3) (vezi Fig. 1.30). Fiecare domeniu de imunoglobulină are o suprastructură β-sheet în care două reziduuri de cisteină sunt legate printr-o legătură disulfurică.
Între cele două domenii constante CH1 și CH2 există o regiune care conține un număr mare de resturi de prolină, care împiedică formarea unei structuri secundare și interacțiunea lanțurilor H învecinate în acest segment. Această regiune balama conferă moleculei de anticorp flexibilitate. Între domeniile variabile ale lanțurilor grele și ușoare există două situsuri identice de legare la antigen (situuri active pentru legare antigene), prin urmare astfel de anticorpi sunt adesea numiți bivalente. Nu întreaga secvență de aminoacizi a regiunilor variabile ale ambelor lanțuri este implicată în legarea antigenului de anticorp, ci doar 20-30 de aminoacizi localizați în regiunile hipervariabile ale fiecărui lanț. Aceste regiuni determină capacitatea unică a fiecărui tip de anticorp de a interacționa cu antigenul complementar corespunzător.
Anticorpii sunt una dintre liniile de apărare ale organismului împotriva organismelor străine invadatoare. Funcționarea lor poate fi împărțită în două etape: prima etapă este recunoașterea și legarea antigenului pe suprafața organismelor străine, ceea ce este posibil datorită prezenței situsurilor de legare a antigenului în structura anticorpului; a doua etapă este inițierea procesului de inactivare și distrugere a antigenului. Specificitatea etapei a doua depinde de clasa de anticorpi. Există cinci clase de lanțuri grele, care diferă unele de altele în structura domeniilor constante: α, δ, ε, γ și μ, conform cărora se disting cinci clase de imunoglobuline: A, D, E, G și M.
Caracteristicile structurale ale lanțurilor grele conferă regiunilor balama și regiunilor C-terminale ale lanțurilor grele o conformație caracteristică fiecărei clase. După ce antigenul se leagă de un anticorp, modificările conformaționale în domeniile constante determină calea pentru îndepărtarea antigenului.
Orez. 1. 30. Structura domeniului IgG
Imunoglobulinele M
Imunoglobulinele M au două forme.
Forma monomerică- clasa I de anticorpi produși prin dezvoltarea limfocitelor B. Ulterior, multe celule B trec la producerea altor clase de anticorpi, dar cu același situs de legare a antigenului. IgM este încorporată în membrană și acționează ca un receptor de recunoaștere a antigenului. Integrarea IgM în membrana celulară este posibilă datorită prezenței a 25 de resturi de aminoacizi hidrofobe în partea de coadă a regiunii.
Forma secretorie a IgM conține cinci subunități monomerice legate între ele prin legături disulfurice și un lanț J polipeptidic suplimentar (Fig. 1.31). Lanțurile grele de monomeri de această formă nu conțin o coadă hidrofobă. Pentamerul are 10 situsuri de legare a antigenului și, prin urmare, este eficient în recunoașterea și îndepărtarea antigenului care intră mai întâi în organism. Forma secretorie a IgM este clasa principală de anticorpi secretați în sânge în timpul răspunsului imun primar. Legarea IgM la un antigen modifică conformația IgM și induce legarea acesteia la prima componentă proteică a sistemului complement (sistemul complement este un set de proteine implicate în distrugerea antigenului) și activarea acestui sistem. Dacă antigenul este situat pe suprafața unui microorganism, sistemul de complement provoacă perturbarea integrității membranei celulare și moartea celulei bacteriene.
Imunoglobulinele G
Cantitativ, această clasă de imunoglobuline predomină în sânge (75% din toate Ig-urile). IgG - monomeri, clasa principală de anticorpi secretați în sânge în timpul unui răspuns imun secundar. După interacțiunea IgG cu antigenele de suprafață ale microorganismelor, complexul antigen-anticorp este capabil să lege și să activeze proteinele sistemului complement sau poate interacționa cu receptorii specifici ai macrofagelor și neutrofilelor. Interacțiunea cu fagocitele conduce
Orez. 1.31. Structura formei secretoare a IgM
la absorbția complexelor antigen-anticorp și distrugerea lor în fagozomii celulari. IgG este singura clasă de anticorpi care sunt capabili să pătrundă în bariera placentară și să ofere protecție intrauterină a fătului împotriva infecțiilor.
Imunoglobulinele A
Clasa principală de anticorpi prezenți în secreții (lapte, saliva, secreții ale tractului respirator și tractului intestinal). IgA este secretată în principal sub formă dimerică, unde monomerii sunt legați unul de celălalt printr-un lanț J suplimentar (Fig. 1.32).
IgA nu interacționează cu sistemul complement și cu celulele fagocitare, dar prin legarea de microorganisme, anticorpii împiedică atașarea lor de celulele epiteliale și pătrunderea în organism.
Imunoglobulinele E
Imunoglobulinele E sunt reprezentate de monomeri în care lanțurile ε grele conțin, ca și lanțurile μ ale imunoglobulinelor M, un domeniu variabil și patru constante. După secreție, IgE se leagă de ea
Orez. 1.32. Structura IgA
Regiunile C-terminale cu receptori corespunzători pe suprafața mastocitelor și bazofilelor. Ca urmare, ei devin receptori pentru antigene de pe suprafața acestor celule (Fig. 1.33).
Orez. 1.33. Interacțiunea IgE cu antigenul de pe suprafața unui mastocit
După ce antigenul se atașează la situsurile corespunzătoare de legare la antigen ale IgE, celulele primesc un semnal pentru a secreta substanțe biologic active (histamină, serotonină), care sunt în mare parte responsabile pentru dezvoltarea reacției inflamatorii și pentru manifestarea reacțiilor alergice, cum ar fi: astm, urticarie, febra fânului.
Imunoglobuline D
Imunoglobulinele D se găsesc în cantități foarte mici în ser; sunt monomeri. Lanțurile grele δ au o variabilă și trei domenii constante. IgD acționează ca receptori pentru limfocitele B; alte funcții sunt încă necunoscute. Interacțiunea antigenelor specifice cu receptorii de pe suprafața limfocitelor B (IgD) duce la transmiterea acestor semnale în celulă și la activarea mecanismelor care asigură proliferarea unei anumite clone de limfocite.
TEMA 1.7. PROPRIETĂȚI FIZICE ȘI CHIMICE ALE PROTEINELOR ȘI METODE DE SEPARARE A LOR
1. Proteinele individuale diferă în proprietăți fizice și chimice:
Forma moleculelor;
Greutate moleculară;
Sarcina totală, a cărei mărime depinde de raportul dintre grupările anionice și cationice ale aminoacizilor;
Raportul dintre radicalii de aminoacizi polari și nepolari de pe suprafața moleculelor;
Grade de rezistență la diferiți agenți de denaturare.
2. Solubilitatea proteinelor depinde asupra proprietăților proteinelor enumerate mai sus, precum și asupra compoziției mediului în care se dizolvă proteina (valori pH, compoziția sării, temperatură, prezența altor substanțe organice care pot interacționa cu proteina). Cantitatea de încărcare a moleculelor de proteine este unul dintre factorii care afectează solubilitatea acestora. Când sarcina din punctul izoelectric se pierde, proteinele se agregează și precipită mai ușor. Acest lucru este tipic în special pentru proteinele denaturate, în care radicalii de aminoacizi hidrofobi apar la suprafață.
Pe suprafața unei molecule de proteine există atât radicali de aminoacizi încărcați pozitiv, cât și negativ. Numărul acestor grupe și, prin urmare, încărcătura totală a proteinelor, depinde de pH-ul mediului, adică. raportul concentrațiilor grupelor H+ - și OH -. Într-un mediu acid O creștere a concentrației de H+ duce la suprimarea disocierii grupărilor carboxil -COO - + H+ > - COOH și la scăderea sarcinii negative a proteinelor. Într-un mediu alcalin, legarea excesului de OH - de către protonii formați în timpul disocierii grupărilor amino -NH 3 + + OH - - NH 2 + H 2 O cu formarea apei, duce la scăderea sarcinii pozitive a proteinelor. . Se numește valoarea pH-ului la care o proteină are o sarcină netă zero punct izoelectric (IEP).În IET, numărul de grupuri încărcate pozitiv și negativ este același, adică. proteina este în stare izoelectrică.
3. Separarea proteinelor individuale. Caracteristicile structurii și funcționării organismului depind de setul de proteine sintetizate în acesta. Studierea structurii și proprietăților proteinelor este imposibilă fără a le izola de celulă și a le purifica de alte proteine și molecule organice. Etape de izolare și purificare a proteinelor individuale:
Distrugerea celulelorţesutul fiind studiat şi obţinerea unui omogenat.
Separarea omogenatului în fracțiuni prin centrifugare, obținând o fracție nucleară, mitocondrială, citosolică sau altă fracție care conține proteina dorită.
Denaturarea termică selectivă- încălzirea pe termen scurt a unei soluții proteice, timp în care unele dintre impuritățile proteice denaturate pot fi îndepărtate (dacă proteina este relativ stabilă la căldură).
Sărare. Diferite proteine precipită la diferite concentrații de sare în soluție. Prin creșterea treptată a concentrației de sare, este posibilă obținerea unui număr de fracții separate cu un conținut predominant de proteine izolate într-una dintre ele. Sulfatul de amoniu este cel mai adesea folosit pentru fracţionarea proteinelor. Proteinele cu cea mai mică solubilitate precipită la concentrații scăzute de sare.
Filtrare pe gel- o metodă de cernere a moleculelor prin granule Sephadex umflate (lanțuri polizaharide tridimensionale de dextran având pori). Viteza cu care proteinele trec printr-o coloană umplută cu Sephadex va depinde de greutatea lor moleculară: cu cât masa moleculelor proteice este mai mică, cu atât ele mai ușor pătrund în granule și rămân acolo mai mult timp; cu cât masa este mai mare, cu atât eluează mai repede din coloană.
Ultracentrifugarea- o metodă care presupune introducerea proteinelor într-un tub de centrifugă în rotorul unei ultracentrifugă. Când rotorul se rotește, viteza de sedimentare a proteinelor este proporțională cu greutatea moleculară a acestora: fracțiile de proteine mai grele sunt situate mai aproape de fundul eprubetei, cele mai ușoare - mai aproape de suprafață.
Electroforeză- o metodă bazată pe diferențele de viteză de mișcare a proteinelor într-un câmp electric. Această valoare este proporțională cu sarcina proteinelor. Electroforeza proteinelor se efectuează pe hârtie (în acest caz, viteza de mișcare a proteinelor este proporțională doar cu sarcina lor) sau într-un gel de poliacrilamidă cu o anumită dimensiune a porilor (viteza de mișcare a proteinelor este proporțională cu sarcina și greutatea moleculară a acestora) .
Cromatografia cu schimb de ioni- o metodă de fracţionare bazată pe legarea grupărilor ionizate de proteine cu grupări încărcate opus de răşini schimbătoare de ioni (materiale polimerice insolubile). Puterea legării proteinei de rășină este proporțională cu sarcina proteinei. Proteinele adsorbite la polimerul schimbător de ioni pot fi spălate cu concentrații crescânde de soluții de NaCl; cu cât este mai mică sarcina de proteine, cu atât este mai mică concentrația de NaCl necesară pentru a spăla proteina legată de grupările ionice ale rășinii.
Cromatografia de afinitate- metoda cea mai specifica de izolare a proteinelor individuale.Un ligand al unei proteine este atasat covalent de un polimer inert. Când o soluție de proteină este trecută printr-o coloană cu un polimer, numai proteina specifică pentru un ligand dat este adsorbită pe coloană datorită legării complementare a proteinei la ligand.
Dializă- o metodă folosită pentru îndepărtarea compușilor cu greutate moleculară mică dintr-o soluție de proteină izolată. Metoda se bazează pe incapacitatea proteinelor de a trece printr-o membrană semi-permeabilă, spre deosebire de substanțele cu greutate moleculară mică. Este folosit pentru a purifica proteinele din impuritățile cu molecularitate scăzută, de exemplu, sărurile după sărare.
TECTURI PENTRU MUNCĂ EXTRAȘCOLARĂ
1. Completați tabelul. 1.4.
Tabelul 1.4. Analiza comparativă a structurii și funcțiilor proteinelor înrudite - mioglobina și hemoglobina
a) amintiți-vă structura centrului activ al lui Mb și Hb. Ce rol joacă radicalii de aminoacizi hidrofobi în formarea centrilor activi ai acestor proteine? Descrieți structura centrului activ al Mb și Hb și mecanismele de atașare a liganzilor la acesta. Ce rol joacă reziduurile His F 8 și His E 7 în funcționarea centrului activ al Mv iHv?
b) ce proprietăți noi are în comparație cu mioglobina monomerică proteina oligomerică strâns înrudită, hemoglobina? Explicați rolul modificărilor cooperante în conformația protomerilor din molecula de hemoglobină, efectul concentrațiilor de CO 2 și protoni asupra afinității hemoglobinei pentru oxigen, precum și rolul 2,3-BPG în reglarea alosterică a funcției Hb .
2. Caracterizați însoțitorii moleculari, acordând atenție relației dintre structura și funcția lor.
3. Ce proteine sunt grupate în familii? Folosind exemplul familiei de imunoglobuline, identificați caracteristicile structurale similare și funcțiile înrudite ale proteinelor acestei familii.
4. Proteinele individuale purificate sunt adesea necesare în scopuri biochimice și medicinale. Explicați pe ce proprietăți fizico-chimice se bazează metodele utilizate pentru separarea și purificarea proteinelor.
SARCINI DE AUTOCONTROL
1. Alege raspunsurile corecte.
Funcțiile hemoglobinei:
A. Transportul O 2 de la plămâni la țesut B. Transportul de H + de la țesuturi la plămâni
B. Menținerea unui pH constant al sângelui D. Transportul CO 2 de la plămâni la țesuturi
D. Transportul CO 2 din țesuturi la plămâni
2. Alege raspunsurile corecte. Ligandα -protomerul Hb este: A. Heme
B. Oxigen
B. CO G. 2,3-BPG
D. β-Protomer
3. Alege raspunsurile corecte.
Hemoglobina spre deosebire de mioglobina:
A. Are o structură cuaternară
B. Structura secundară este reprezentată doar de elice α
B. Aparține proteinelor complexe
D. Interacționează cu un ligand alosteric D. Legat covalent de hem
4. Alege raspunsurile corecte.
Afinitatea Hb pentru O2 scade:
A. Când se adaugă o moleculă de O 2 B. Când o moleculă de O 2 este îndepărtată
B. Când interacționați cu 2,3-BPG
D. Când este atașat de protomeri H + D. Când concentrația de 2,3-BPG scade
5. Meci.
Tipurile HB se caracterizează prin:
A. În formă deoxi formează agregate fibrilare B. Conține două lanțuri α și două δ
B. Forma predominantă de Hb în eritrocitele adulte D. Conține hem cu Fe+ 3 în centrul activ
D. Conține două lanțuri α și două γ 1. HbA 2.
6. Meci.
Liganzii Hb:
A. Se leagă de Hb în centrul alosteric
B. Are o afinitate foarte mare pentru situsul activ al Hb
B. Prin unire, crește afinitatea Hb pentru O 2 G. Oxidează Fe+ 2 la Fe+ 3
D. Formează o legătură covalentă cu hisF8
7. Alege raspunsurile corecte.
Însoțitori:
A. Proteine prezente în toate părțile celulei
B. Sinteza crește sub stres
B. Participa la hidroliza proteinelor denaturate
D. Participa la menținerea conformației native a proteinelor
D. Ele creează organite în care se formează conformația proteinelor.
8. Meci. Imunoglobuline:
A. Forma secretorie este pentamerică.
B. Clasa Ig care pătrunde în bariera placentară
B. Ig - receptor mastocitar
D. Clasa principală de Ig prezentă în secrețiile celulelor epiteliale. D. Receptorul limfocitelor B, a cărui activare asigură proliferarea celulară
9. Alege raspunsurile corecte.
Imunoglobulinele E:
A. Produse de macrofage B. Au lanțuri ε grele.
B. Înglobat în membrana limfocitelor T
D. Acționează ca receptori de antigen de membrană pe mastocite și bazofile
D. Responsabil pentru reacțiile alergice
10. Alege raspunsurile corecte.
Metoda de separare a proteinelor se bazează pe diferențele de greutate moleculară a acestora:
A. Filtrare pe gel
B. Ultracentrifugarea
B. Electroforeza pe gel de poliacrilamidă D. Cromatografia de schimb ionic
D. Cromatografia de afinitate
11. Alege răspunsul corect.
Metoda de separare a proteinelor se bazează pe diferențele de solubilitate a acestora în apă:
A. Filtrare pe gel B. Sărare
B. Cromatografia de schimb ionic D. Cromatografia de afinitate
D. Electroforeza în gel de poliacrilamidă
STANDARDE DE RĂSPUNSURI LA „SARCINI DE AUTOCONTROL”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
TERMENI ȘI CONCEPTE DE BAZĂ
1. Proteine oligomerice, protomer, structura cuaternară a proteinelor
2. Modificări cooperante în conformația protomerului
3. Efectul Bohr
4. Reglarea alosterică a funcțiilor proteice, centru alosteric și efector alosteric
5. Chaperone moleculare, proteine de șoc termic
6. Familii de proteine (serin proteaze, imunoglobuline)
7. Relația IgM-, G-, E-, A-structură-funcție
8. Sarcina totală a proteinelor, punctul izoelectric al proteinelor
9. Electroforeza
10. Sărare
11. Filtrare pe gel
12. Cromatografia de schimb ionic
13. Ultracentrifugarea
14. Cromatografia de afinitate
15. Electroforeza proteinelor plasmatice sanguine
SARCINI PENTRU MUNCĂ LA CLASĂ
1. Comparați dependențele gradelor de saturație ale hemoglobinei (Hb) și mioglobinei (Mb) cu oxigenul de presiunea sa parțială în țesuturi
Orez. 1.34. Dependenţa de saturaţie Mv şiNHoxigen din presiunea sa parțială
Vă rugăm să rețineți că forma curbelor de saturație a oxigenului proteinei este diferită: pentru mioglobină - o hiperbolă, pentru hemoglobină - o formă sigmoidă.
1. comparați valorile presiunii parțiale a oxigenului la care Mb și Hb sunt saturate cu O 2 cu 50%. Care dintre aceste proteine are o afinitate mai mare pentru O2?
2. Ce caracteristici structurale ale Mb determină afinitatea sa mare pentru O 2?
3. Ce caracteristici structurale ale HB îi permit să elibereze O2 în capilarele țesuturilor în repaus (la o presiune parțială relativ mare a O2) și să mărească brusc această eliberare în mușchii care lucrează? Ce proprietate a proteinelor oligomerice oferă acest efect?
4. Calculați ce cantitate de O 2 (în %) o dă hemoglobina oxigenată mușchilor care se odihnesc și lucrează?
5. trageți concluzii despre relația dintre structura unei proteine și funcția acesteia.
2. Cantitatea de oxigen eliberată de hemoglobină în capilare depinde de intensitatea proceselor catabolice din țesuturi (efect Bohr). Cum modificările metabolismului tisular reglează afinitatea Hb pentru O2? Efectul CO2 și H+ asupra afinității Hb pentru O2
1. descrie efectul Bohr.
2. în ce direcție se desfășoară procesul prezentat în diagramă:
a) în capilarele plămânilor;
b) în capilarele tisulare?
3. Care este semnificația fiziologică a efectului Bohr?
4. De ce interacțiunea Hb cu H+ la locuri îndepărtate de hem modifică afinitatea proteinei pentru O 2?
3. Afinitatea Hb pentru O2 depinde de concentrația ligandului său - 2,3-bisfosfoglicerat, care este un regulator alosteric al afinității Hb pentru O2. De ce interacțiunea ligandului la un loc îndepărtat de locul activ afectează funcția proteinei? Cum reglează 2,3-BPG afinitatea Hb pentru O2? Pentru a rezolva problema, răspunde la următoarele întrebări:
1. unde și din ce se sintetizează 2,3-bisfosfogliceratul (2,3-BPG)? Scrieți formula acesteia, indicați sarcina acestei molecule.
2. Cu ce formă de hemoglobină (oxi sau deoxi) interacționează BPG și de ce? În ce parte a moleculei de Hb are loc interacțiunea?
3. în ce direcție are loc procesul prezentat în diagramă?
a) în capilarele tisulare;
b) în capilarele plămânilor?
4. unde concentraţia complexului ar trebui să fie mai mare
Nv-2,3-BFG:
a) în capilarele mușchilor în repaus,
b) în capilarele mușchilor care lucrează (cu condiția aceeași concentrație de BPG în eritrocite)?
5. Cum se va schimba afinitatea HB pentru oxigen atunci când o persoană se adaptează la condițiile de altitudine mare, dacă crește concentrația de BPG în eritrocite? Care este semnificația fiziologică a acestui fenomen?
4. Distrugerea 2,3-BPG în timpul depozitării sângelui conservat afectează funcțiile HB. Cum se va schimba afinitatea HB pentru O 2 în sângele conservat dacă concentrația de 2,3-BPG în eritrocite poate scădea de la 8 la 0,5 mmol/l. Este posibil să se transfuzeze un astfel de sânge pacienților grav bolnavi dacă concentrația de 2,3-BPG este restabilită nu mai devreme de trei zile? Este posibil să restabiliți funcțiile celulelor roșii din sânge prin adăugarea de 2,3-BPG în sânge?
5. Amintiți-vă structura celor mai simple molecule de imunoglobuline. Ce rol joacă imunoglobulinele în sistemul imunitar? De ce Ig-urile sunt adesea numite bivalente? Cum se raportează structura Ig-urilor cu funcția lor? (Descrieți folosind un exemplu de clasă de imunoglobuline.)
Proprietățile fizico-chimice ale proteinelor și metodele de separare a acestora.
6. Cum afectează încărcătura netă a unei proteine solubilitatea acesteia?
a) determinați sarcina totală a peptidei la pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) cum se va schimba sarcina acestei peptide la pH >7, pH<7, рН <<7?
c) care este punctul izoelectric al unei proteine (IEP) și în ce mediu se află?
IET al acestei peptide?
d) la ce valoare pH se va observa cea mai mică solubilitate a acestei peptide.
7. De ce laptele acru, spre deosebire de laptele proaspăt, se „coagulează” atunci când este fiert (adică precipită cazeina din proteinele din lapte)? În laptele proaspăt, moleculele de cazeină au o sarcină negativă.
8. Filtrarea pe gel este utilizată pentru a separa proteinele individuale. Un amestec care conține proteine A, B, C cu greutăți moleculare egale cu 160.000, 80.000 și, respectiv, 60.000, a fost analizat prin filtrare pe gel (Fig. 1.35). Granulele de gel umflate sunt permeabile la proteinele cu o greutate moleculară mai mică de 70 000. Ce principiu stă la baza acestei metode de separare? Care grafic reflectă corect rezultatele fracționării? Indicați ordinea în care proteinele A, B și C sunt eliberate din coloană.
Orez. 1.35. Utilizarea gelului de filtrare pentru separarea proteinelor
9. În fig. 1.36, A prezintă o diagramă a electroforezei pe hârtie a proteinelor din serul sanguin al unei persoane sănătoase. Cantitățile relative de fracții proteice obținute prin această metodă sunt: albumine 54-58%, α1-globuline 6-7%, α2-globuline 8-9%, β-globuline 13%, y-globuline 11-12%.
Orez. 1.36 Electroforeza pe hârtie a proteinelor plasmatice ale unei persoane sănătoase (A) și ale unui pacient (B)
I - y-globuline; II - β-globuline; III -α 2 -globulina; IV -α 2 -globulina; V - albumine
Multe boli sunt însoțite de modificări cantitative ale compoziției proteinelor serice (disproteinemie). Natura acestor modificări este luată în considerare atunci când se pune un diagnostic și se evaluează severitatea și stadiul bolii.
Folosind datele din tabel. 1.5, faceți o ghicire despre boală, care se caracterizează prin profilul electroforetic prezentat în Fig. 1.36.
Tabelul 1.5. Modificări ale concentrației proteinelor serice în patologie


